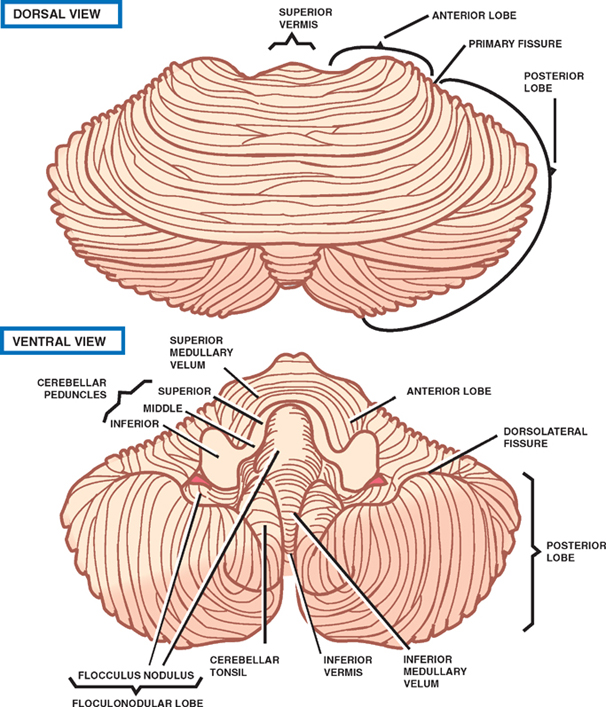
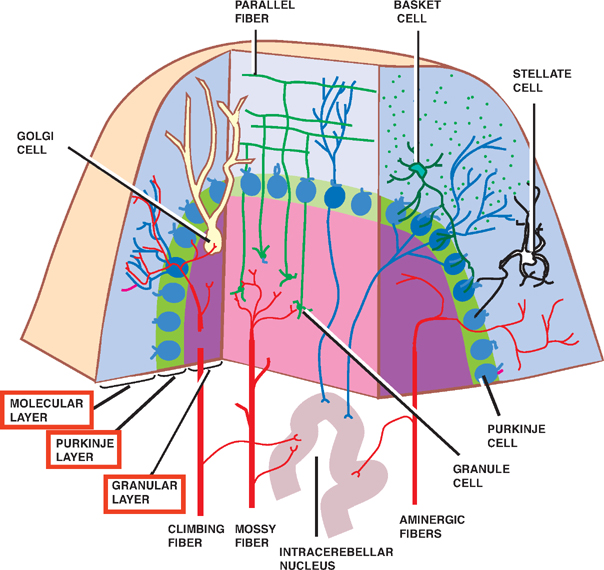
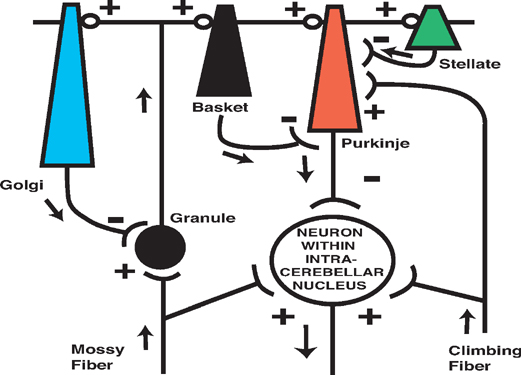
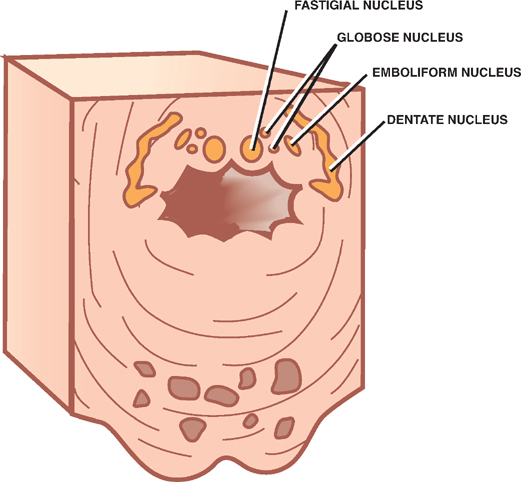
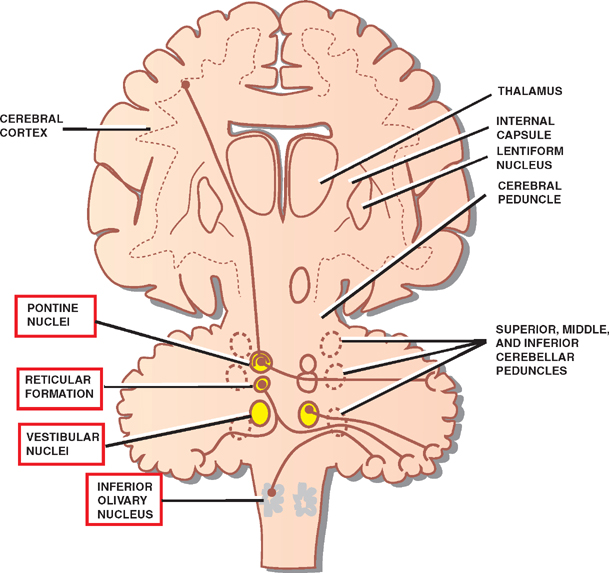
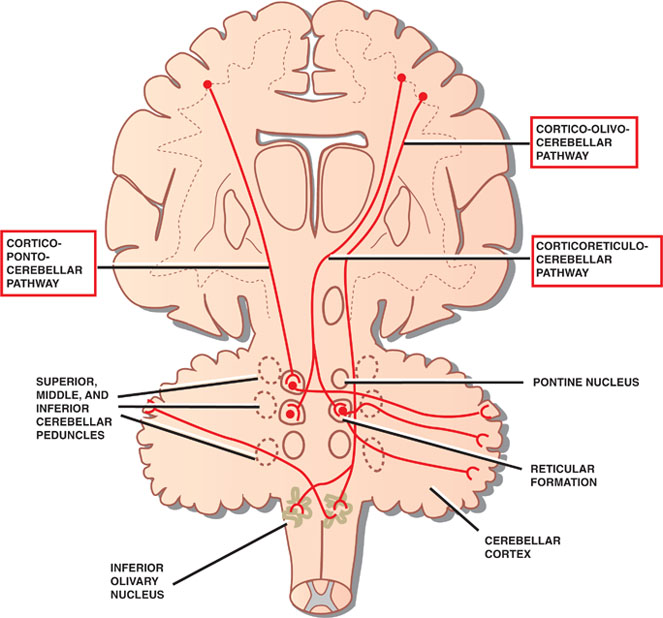
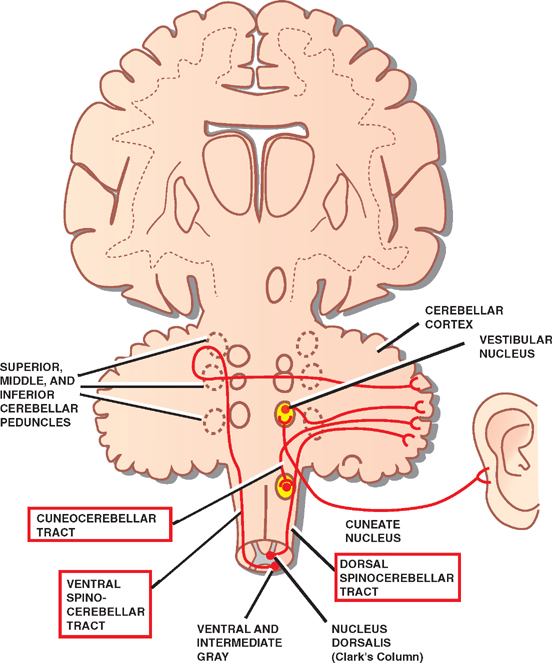
8 Cerebellum See Fig. 8.1. The cerebellum is composed of two cerebellar hemispheres joined by a median vermis. The superior vermis is confluent with the hemispheres, whereas the inferior vermis is a well-delineated structure that is located in a deep depression in the midline (the vallecula). The cerebellum is divided into three lobes: the anterior lobe, the posterior lobe, and the flocculonodular lobe. The anterior lobe (paleocerebellum) constitutes the rostral portion of the rostral cerebellar surface; the floc-culonodular lobe (archicerebellum) constitutes the rostral portion of the caudal cerebellar surface; and the posterior lobe (neocerebellum) makes up the remainder of the cerebellum on both surfaces. Two major fissures of the cerebellum are identified: the primary fissure separates the anterior lobe from the posterior lobe on the superior surface, and the dorsolateral fissure separates the posterior lobe from the floc-culonodular lobe on the inferior surface. The rostral portion of the roof of the fourth ventricle is formed by the superior cerebellar peduncles. The caudal aspect of the roof of the fourth ventricle is formed by the inferior medullary velum, which frequently adheres to the inferior vermis. The foramen of Magendie is a median aperture, which constitutes an opening in the inferior medullary velum that connects the fourth ventricle with the cisterna magna. The three cerebellar peduncles are attached to the cerebellum in the interval between the anterior and the flocculonodular lobes. Fig. 8.1 External appearance of cerebellum. See Fig. 8.2. The cerebellar cortex is divided into three layers: a superficial molecular layer, a middle Purkinje layer, and a deep granular layer. These layers are collectively composed of five cell types: stellate, basket, Purkinje, Golgi, and granule cells. The molecular layer is so-called because of its punctuated, sparsely populated appearance. It is largely a synaptic layer that contains dendrites of Purkinje cells and axons of granule cells. Scattered stellate and basket cells are also present. The Purkinje cell layer consists of a single row of Purkinje cell bodies; the granule cell layer consists of densely packed neurons that send axonal projections into the molecular layer. The granule cell layer is composed of granule and Golgi cells. An embryonic cerebellar layer, the external granule cell layer, is present during the prenatal and early postnatal periods but is completely gone by the first year of life. This layer is superficial to the molecular layer, and its cells are thought to be the cell of origin of medullo-blastomas. Fig. 8.2 Layers of the cerebellum. See Fig. 8.3. The intrinsic circuitry of the cerebellar cortex is primarily composed of two interrelated circuits, which receive input from two afferent nerve fibers, the climbing fibers and the mossy fibers. The input from these fiber systems is received by the Purkinje cells (directly and indirectly), which in turn project to the neurons in the deep cerebellar nuclei. Climbing fibers, most of which originate in the inferior olive, make direct contacts with the dendrites of a limited number of Purkinje cells. By contrast, mossy fibers, which are derived from a variety of sources, indirectly influence a large number of Purkinje cells. To accomplish this, the mossy fibers branch extensively in the granule cell layer, where they establish synaptic contacts with the granule cells. Rosettes are the sites of synapses between mossy fibers and the clawlike terminals of granule cell dendrites. Mossy fiber rosettes also establish synapses with Golgi type II cell bodies. The granule cells project superficially to the molecular layer, where they bifurcate in a T-shaped manner to form the so-called parallel fibers. Each parallel fiber establishes contacts with a large number of Purkinje cell dendrites. Thus, the climbing fibers exert a powerful influence on a few specific Purkinje cells, whereas the mossy fibers modulate a large number of Purkinje cells through a relay in the granular layer. In other words, a single climbing fiber may stimulate a Purkinje cell in an all-or-none phenomenon, whereas many mossy fiber discharges are required to stimulate a Purkinje cell. A third category of afferent axons, the aminergic fibers, are distinct both from climbing and mossy fibers in that they contain biogenic amines. These fibers, which show a widespread distribution in the cerebellar cortex, are divided into two different types: serotonin-containing axons that originate in the raphe nuclei of the brain-stem, and norepinephrine-containing axons that originate in the locus ceruleus. These circuits involving climbing fibers, mossy fibers, and aminergic fibers are modified by intracortical circuits formed by three types of interneurons that appear to function as modulators of Purkinje cell activity. These are the Golgi, basket, and stellate cells. Like the Purkinje cells, these interneurons are inhibitory in nature, and they contain the neurotransmitter gamma-aminobutyric acid (GABA). Thus, four out of the five cells in the cerebellar cortex are GABA-containing inhibitory neurons. Only the granule cells and the afferent fiber system are excitatory. The overall circuit in the cerebellar cortex may be summarized as follows. Excitatory input to the cerebellar cortex is primarily derived from the mossy fibers and the climbing fibers. This excitatory input is received by the Purkinje cells (directly and indirectly), which are responsible, in turn, for the entire (inhibitory) output of the cerebellar cortex. The excitatory input to the Purkinje cells is further modified by the inhibitory influences of the modulating interneurons. The Purkinje cells project into the deep cerebellar nuclei. Fig. 8.3 Intrinsic circuitry. See Fig. 8.4. Four pairs of nuclei are located in the white matter of the cerebellum. From medial to lateral on each side of the midline, they are the fastigial, globose, emboliform, and dentate nuclei. The globose and the emboliform nuclei are collectively known as the interposed nuclei because they are interposed between the fastigial and dentate nuclei. The input to the cerebellar nuclei is derived from two sources: (1) excitatory input is derived from fibers that originate in cells that lie outside the cerebellum, and (2) inhibitory input is derived from fibers that arise from the Purkinje cells of the cortex. Cells outside the cerebellum that send afferents directly to the cerebellar nuclei include pontocerebellar, spinocerebellar, and olivocerebellar fibers, most of which give collaterals to the cerebellar nuclei, then continue on to the cerebellar cortex. The cells that constitute the intracerebellar nuclei act to modify muscular activity through the motor control areas of the brainstem and cerebral cortex. Efferents from the fastigial nucleus project to the brainstem through the inferior cerebellar peduncle, whereas efferents from the other nuclei are projected to the brainstem and cerebral cortex (with relays in the thalamus) through the superior cerebellar peduncle. Fig. 8.4 Deep cerebellar nuclei. See Fig. 8.5. Cerebellar afferents are derived from three main sources: the cerebral cortex, the spinal cord, and the vestibular nerve. A small number of afferents originate in the red nucleus and the tectum. Fig. 8.5 Afferent cerebellar connections. See Fig. 8.6. Before reaching the cerebellum, cortical projections establish synaptic contacts with three brainstem structures: (1) the pontine nuclei, (2) the inferior olivary nucleus, and (3) the reticular formation. The corticopontocerebellar pathway originates from a large area of the cerebral cortex, descends through the corona radiata and the internal capsule, and terminates in the pontine nuclei. The cells of the pontine nuclei give rise to mossy fibers that cross the midline to reach the opposite cerebellar hemisphere via the middle cerebellar peduncle. The cortico-olivocerebellar pathway also originates from a large area of the cerebral cortex, descends through the corona radiata and the internal capsule, and terminates bilaterally in the inferior olivary nuclei. The cells of the inferior olivary nuclei give rise to climbing fibers that cross the midline to enter the opposite cerebellar hemisphere via the inferior cerebellar peduncle. The corticoreticulocerebellar pathway originates from a large area of the cerebral cortex, descends through the corona radiata and the internal capsule, and terminates bilaterally in the reticular formation of the pons and medulla. The cells of the reticular formation give rise to mossy fibers that enter the ipsilateral cerebellar hemisphere via the inferior and the middle cerebellar peduncles. See Fig. 8.7, p. 288. Spinal cord projections to the cerebellum are carried in three spinocerebellar pathways: (1) the ventral spino-cerebellar tract, (2) the dorsal spinocerebellar tract, and (3) the cuneocerebellar tract. Fig. 8.6 Afferent cerebellar connections from cortex. See Fig. 8.7. The vestibular nerve gives rise to afferent fibers that terminate in the vestibular nuclei of the brainstem. The neurons of the vestibular nuclei in turn give rise to mossy fibers that pass through the inferior cerebellar peduncle to enter the ipsilateral flocculonodular lobe on the same side.
External Appearance
Cerebellar Cortex
Cortical Layers
Intrinsic Circuitry
Deep Cerebellar Nuclei
Cerebellar Connections
Afferent Connections
Cerebral Cortex
Spinal Cord
Vestibular Nerve
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


