2
Microsurgical Anatomy of the Posterior Cranial Fossa
The posterior cranial fossa is unique in the cranial cavity, as it is the largest of the three cranial fossae and has the most complex intracranial anatomy. It harbors the parts of the brain that control respiration, cardiac cycle, consciousness, and balance. It is the conduit of fibers of practically all important motor and sensory pathways. Here lies the conduit in which the cerebrospinal fluid (CSF) emerges from the ventricular system into the subarachnoid space. Moreover, the complex arterial relationships of the vertebrobasilar system make this region even more complex. In addition to discussing surgical anatomy, this chapter highlights the anatomy of common surgical approaches to this region, and provides illustrations of radiologic anatomy and some endoscopic anatomy (Figs. 2.1, 2.2, 2.3, and 2.4), which is essential for a neurosurgeon in practice or in training.
♦ Panoramic View of the Posterior Fossa
Superiorly, the posterior fossa starts from the tentorial hiatus and extends down to the foramen magnum. It is encased by occipital, temporal, and sphenoid bones, with a small contribution from parietal bone. In front of it is the dorsum sellae and the clival part of the occipital bone; behind it is the squamous part of the occipital bone, and on both sides are the petrous and mastoid part of the temporal bone along with the lateral part of the squamous part of the occipital bone. The upper peripheral part is bounded by the sulcus of the lateral sinus starting from the internal occipital protuberance and extending anterolaterally to the posterior part of the petrous ridge. The boundary continues along the petrous ridge to end at the petroclinoid junction (Fig. 2.5). We will discuss the bony architecture as well as the neural structures and vessels.
♦ Bony Anatomy
Foramen Magnum and Its Adjacent Region
When the posterior cranial fossa is viewed from above or below in a dry skull specimen, the foramen magnum is the most prominent landmark that comes into view. This ovalshaped foramen is wider posteriorly, and this posterior parts transmits the medulla. The narrower anterior part sits over the odontoid peg.
The posterior part of the foramen magnum is contributed to by the squamous part of the occipital bone, which is an internally concave plate. This part not only forms the posterior boundary but also continues above the foramen magnum. The convex external surface has several prominences on which muscles of the neck attach; these muscles include the sternocleidomastoid, trapezius, splenius capitis, semispinalis capitis, rectus capitis posterior major, and superior oblique. The largest prominence, the external occipital protuberance or inion, is located about a centimeter below the apex of the internal occipital protuberance and is slightly below the torcular Herophili. This is an important landmark for performing a craniotomy in the posterior fossa exposure. The inner surface has the internal occipital protuberance near its center. The internal surface is divided into four somewhat unequal parts by bony ridges. The lower two parts are bounded medially by the internal occipital crest (a prominent ridge descending from the internal occipital protuberance, which gives attachment to falx cerebelli) and above by the paired sulci for lateral sinuses that extend laterally from the internal occipital protuberance. The sulcus for the right lateral sinus is usually larger (Fig. 2.5). The internal occipital crest bifurcates above the foramen magnum to form paired lower limbs, between which there is a depression called the vermian fossa, which is occupied by the lower end of the vermis of the cerebellum.
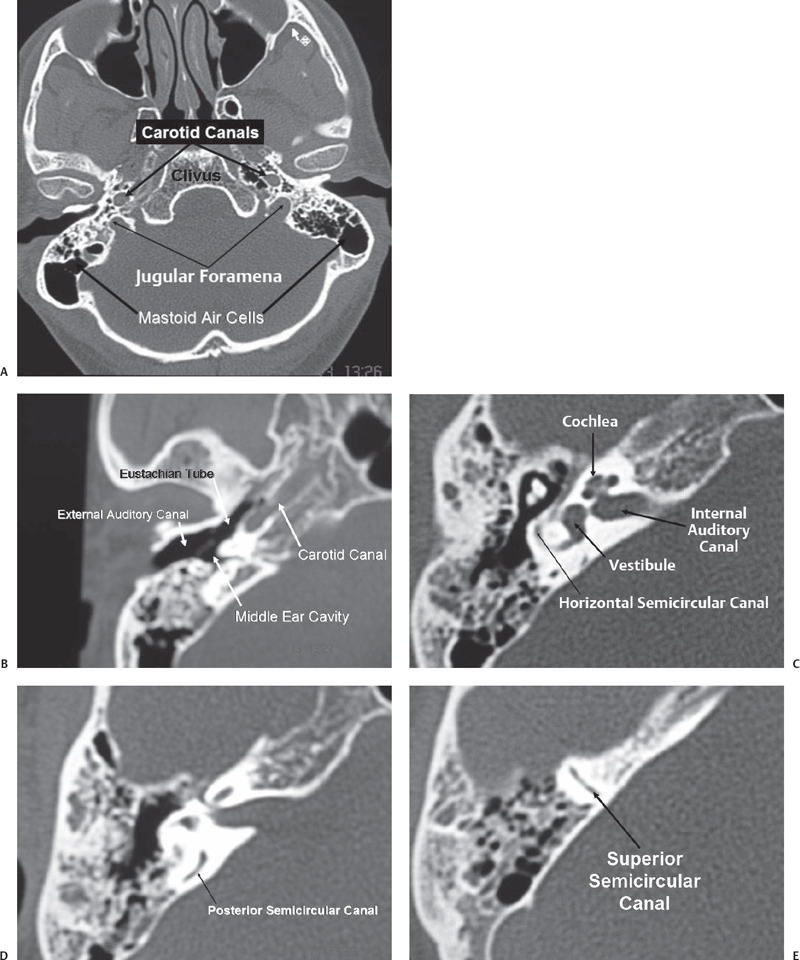
Fig. 2.1 Bone window of computed tomography (CT) scan showing the crucial anatomy of the posterior fossa. (A) Axial section of the posterior fossa showing the relationship of the carotid canal and the jugular foramina. (B) Section through the temporal bone showing the external ear, middle ear, and eustachian tube. Note the relationship of the eustachian tube and the carotid canal. (C) Another section through the temporal bone showing the bony labyrinth and the internal auditory canal. (D) A section through the posterior semicircular canal. Note its proximity to the posterior surface of the petrous bone. (E) A section through the superior semicircular canal.
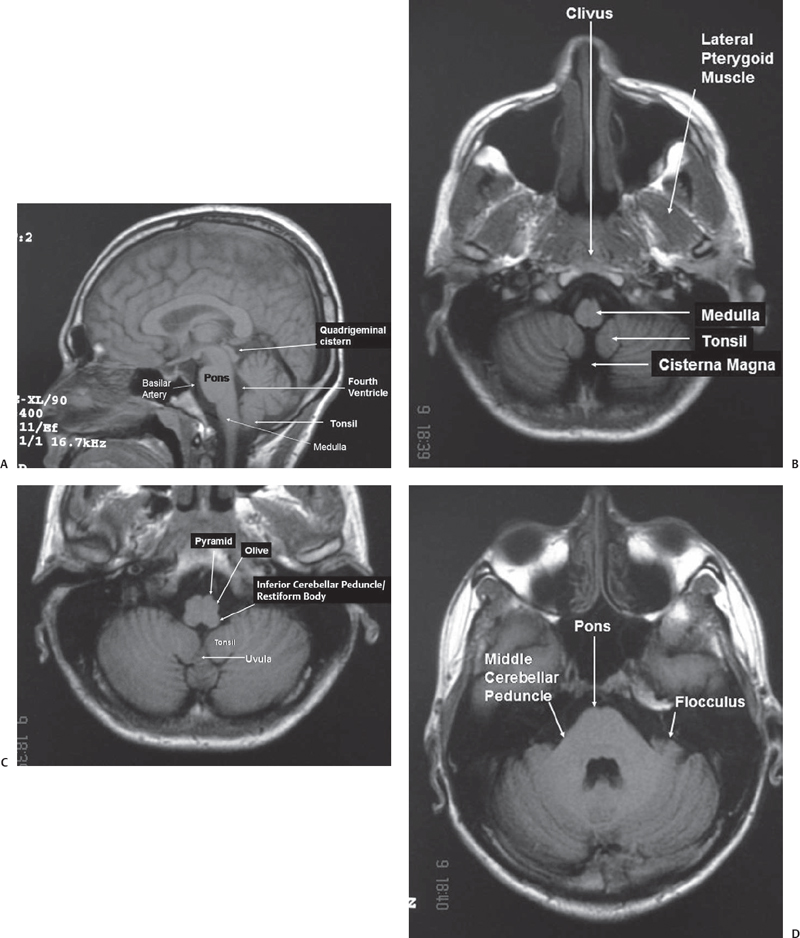
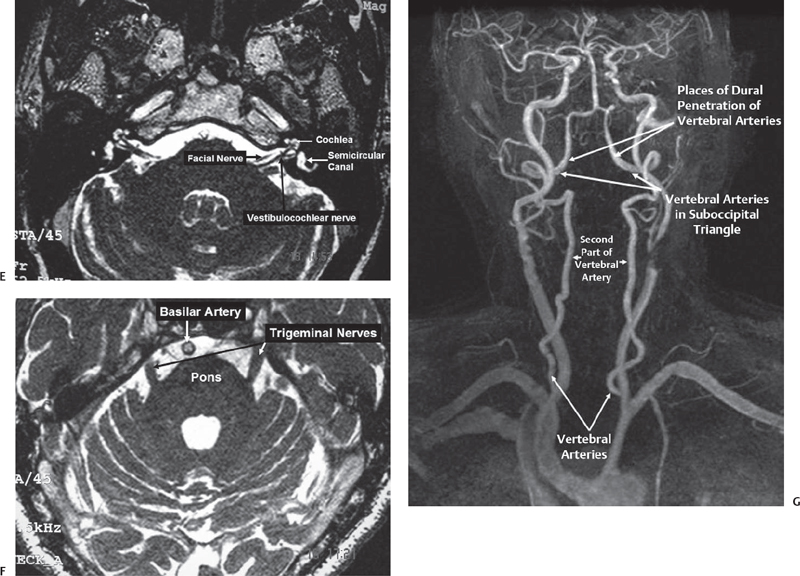
Fig. 2.2 Magnetic resonance imaging (MRI) scan showing important anatomy of the posterior fossa. (A) Sagittal T1 MRI scan showing the brainstem, fourth ventricle, and cerebellar tonsil. (B) Axial section through lower medulla showing the cisterna magna and the tonsil of the cerebellum. (C) Axial section a little above the previous section showing parts of the medulla, uvula, and tonsil of cerebellum. (D) Axial section through the midpons. (E) High-resolution MRI showing exquisite detail of the anatomy of the acousticofacial bundle and labyrinth. (F) High-resolution MRI showing a section through the midpons. (G) MR angiogram showing the vertebrobasilar system.
The anterior part of the foramen magnum is formed by the basilar part of the occipital bone or the clivus, which is a thick quadrangular plate of bone that extends forward and upward at an angle of 45 degrees from the foramen magnum. If traced upward, the spheno-occipital synchondrosis (where the occipital bone joins the sphenoid bone) is found just below the dorsum sellae. The dorsal part of the clivus is concave from side to side, and laterally it is bounded by the petroclival fissure (which separates the clivus from the petrous temporal bone) on both sides (Fig. 2.6). The petroclival fissure harbors the inferior petrosal sinus, and if traced back, it ends in the jugular foramen. On the ventral surface of the clivus just in front of the foramen magnum there is a small elevation called the pharyngeal tubercle (Fig. 2.6), which gives attachment to the pharyngeal raphe.
Laterally, the foramen magnum is bounded by the condylar parts of the occipital bone. The most prominent part of the condylar part is the paired occipital condyles, which are oval in shape and convex downward; they face downward and laterally, with the long axes in the anteromedial direction, and they are located lateral to the anterior half of the foramen magnum and protrude from the condylar part of the occipital bone. They articulate with the lateral masses of the atlas. On the external surface behind the condyle there is the condylar fossa (Fig. 2.7), a depression that is often perforated to form the condylar canal transmitting the emissary veins. During the transcondylar approach, these veins can cause troublesome bleeding. From the posterior half of the condyle, the jugular process, a quadrilateral plate of bone extends laterally to form the posterior border of the jugular foramen. On the medial side of each condyle there is a tubercle, which gives attachment to the alar ligament of the odontoid process. The jugular tubercle, an oval prominence on the intracranial surface of the condylar part, sits just above the hypoglossal canal and medial to the lower extent of the petroclival fissure (Fig. 2.6).
Jugular Foramen Region
The jugular foramen, sitting at the posterior end of the petroclival fissure, is situated lateral and slightly superior to the anterior half of the occipital condyle (Fig. 2.8). It is bordered posteriorly by the jugular process of the occipital bone, and anterosuperiorly by the jugular fossa of the petrous temporal bone.1 It can be considered a hiatus between the temporal and occipital bones. The right foramen is larger than the left. The long axis of the foramen is directed anteromedially, so that its anterolateral margin is formed by the temporal bone, and the posteromedial margin is formed by the occipital bone. It is directed anteromedially and downward from the intracranial aspect. The foramen cannot be seen when the base of the skull is viewed directly from above or below. It has a large oval component laterally, where it receives the drainage of the sigmoid sinus, and a smaller medial part or petrosal part, which receives the drainage of the inferior petrosal sinus (Fig. 2.8A). If the foramen is viewed from below, the part of the jugular foramen that is seen is the part on which sits the jugular bulb. The intrajugular processes on the opposing surfaces of the occipital and temporal bones are connected by a fibrous strand, called the intrajugular septum. This septum, which can sometimes be bony, separates the petrosal and sigmoid part of the foramen. The intrajugular ridge extends forward from the intrajugular process of the temporal bone along the medial edge of the jugular bulb (Fig. 2.9B). The glossopharyngeal nerve courses along the medial edge of this ridge. Sometimes a canal is formed through which the glossopharyngeal nerve courses.
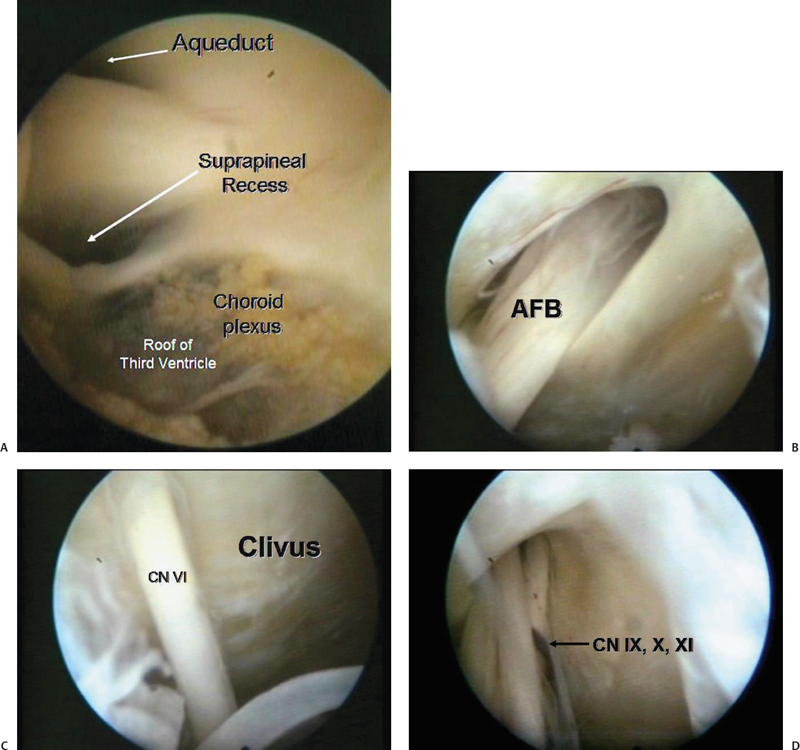
Fig. 2.3 Demonstration of endoscopic anatomy in a cadaver after a third ventriculostomy. The endoscope was navigated through the prepontine cistern and below. (A) Oculomotor nerve on the left side is seen. (B) Acousticofacial bundle (AFB) is seen. (C) Abducens is seen entering Dorello’s canal. CN, cranial nerve. (D) Lower cranial nerves.
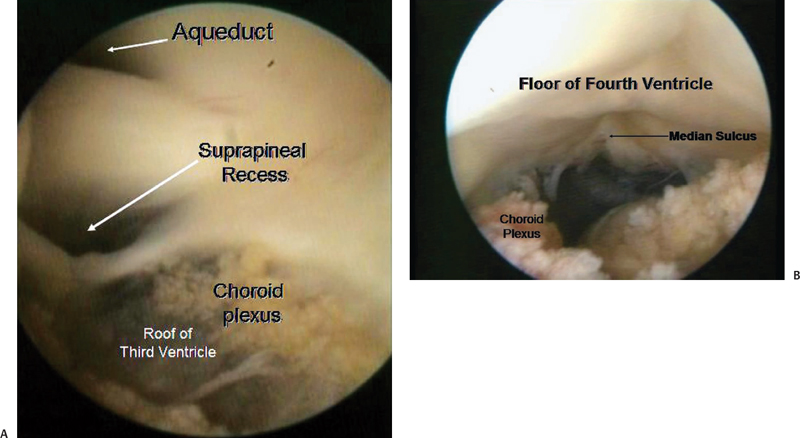
Fig. 2.4 The endoscope was navigated back from the third ventricle to guide through the aqueduct into the fourth ventricle. (A) The roof of third ventricle is clearly seen, along with the aqueduct and the suprapineal recess. (B) Entry into the fourth ventricle.

Fig. 2.5 A panoramic view of the bony posterior cranial fossa from above.
The sigmoid sinus courses forward into the sigmoid portion of the jugular foramen. In this foramen there is a high domed recess, called the jugular fossa, which forms the roof over the jugular bulb and is usually larger on the right side due to the larger size of the jugular bulb on the right side. On the medial side of the intrajugular process of the temporal bone, a small triangular recess, called the pyramidal fossa, extends forward along the anterior wall of the petrosal part of the foramen. The external aperture of the cochlear canaliculus, housing the perilymphatic duct and a tubular prolongation of the dura mater, opens into the anterior apex of the pyramidal fossa (Fig. 2.9A). The glossopharyngeal nerve enters this fossa below the point at which the cochlear aqueduct joins its apex.
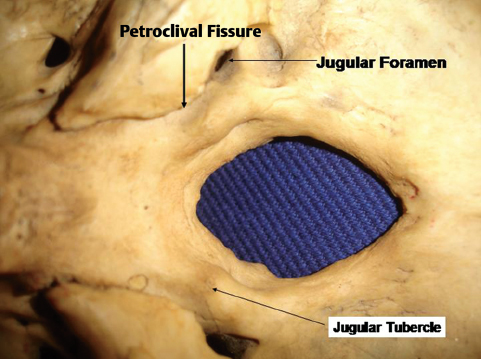
Fig. 2.6 Foramen magnum and the adjacent region from above.
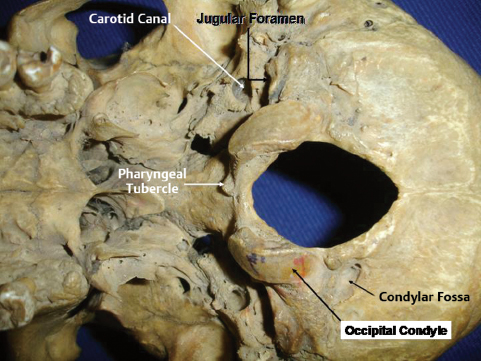
Fig. 2.7 Base of the skull from below.
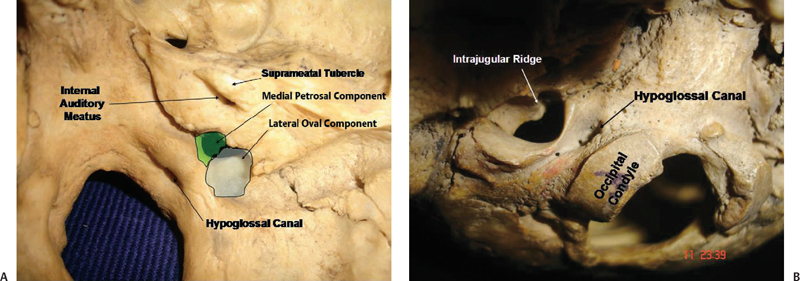
Fig. 2.8 View of the jugular foramen of the right side from inside (A) and outside (B).
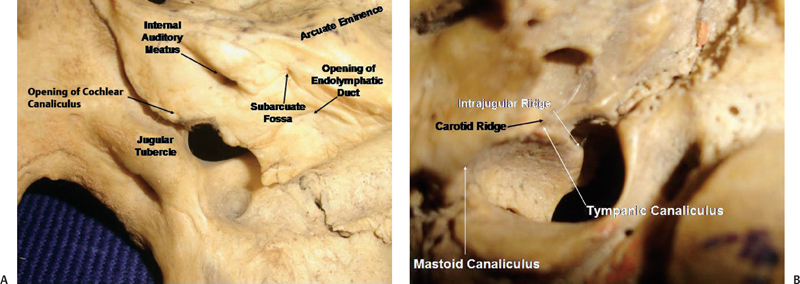
Fig. 2.9 Close-up view of the jugular foramen region from inside (A) and outside (B).
The jugular process of the occipital bone, forming the posteromedial wall of the foramen, extends laterally from the area above the posterior half of the occipital condyle. It is penetrated by the hypoglossal canal. An oval prominence, the jugular tubercle, is found on the superior surface of the intrajugular process and is exactly above the hypoglossal canal (Fig. 2.9A). Occasionally, the jugular tubercle is marked by a shallow furrow indicating the passage of the glossopharyngeal, vagus, and accessory nerves.
The hypoglossal canal passes through the condylar part of the occipital bone and is located medial to the jugular foramen just below the jugular tubercle and approximately 5 mm inferomedial to the petrosal part of the jugular foramen (Figs. 2.8A and 2.9A).
Just inside the lateral edge of the jugular foramen there is the mastoid canaliculus, through which the auricular branch of the vagus (Arnold’s nerve) enters the mastoid. The nerve exits the bone in the inferolateral part of the tympanomastoid suture. A small canal, the tympanic canaliculus, is located at the site where the intrajugular ridge of the temporal bone meets the carotid ridge, which separates the carotid canal and jugular foramen, and is directed upward. The tympanic branch of the inferior glossopharyngeal ganglion (Jacobson’s nerve) courses through this area to the tympanic cavity (Fig. 2.9B).
Petrous Temporal Bone and the Adjoining Region
The petrous temporal bone is probably the most important bone for neurosurgeons. Its intricate anatomy must be learned in detail before venturing into surgery of this region. As many as seven cranial nerves are related to this bone. The internal carotid artery has a sizable length in this bone. The petrous part of the temporal bone is a pyramid-shaped with a base, an apex, three surfaces, and margins, and is wedged between the sphenoid and occipital bones. The anterior or superior surface faces the middle cranial fossa and is not discussed here.
The posterior surface of the petrous bone is continuous with the mastoid surface and faces the cerebellopontine angle region. The internal auditory meatus (IAM) is located midway between the base and apex on the posterior surface (Figs. 2.8A and 2.9A). The lateral end of the internal auditory ends in the meatal fundus, which is divided into superior and inferior parts by the transverse crest. The superior part is occupied by the facial nerve anteriorly and the superior vestibular nerve posteriorly. The inferior part is occupied by the cochlear nerve anteriorly and inferiorly. The superior part is incompletely divided by another bony bar, called the vertical crest or Bill’s bar (Fig. 2.10A), which separates the anteriorly located facial canal from the posteriorly located superior vestibular nerve.2 Posterior to the entry of the inferior vestibular nerve into the semicircular canal, in the posteroinferior corner of the meatal fundus, there is the singular foramen for the singular branch of the inferior vestibular nerve, innervating the posterior ampullae. Just lateral to the porus acusticus on the posterior surface of the petrous bone is the site of a small bony opening, called the subarcuate fossa, which gives passage to the subarcuate artery, a branch of the anterior inferior cerebellar artery (AICA), which usually ends blindly in the region of the superior semicircular canal. The opening for the vestibular aqueduct transmitting the endolymphatic duct, which opens below into the endolymphatic sac in between the dural layers, is inferolateral to the porus (Fig. 2.9A). Inferior to the porus, at the anteromedial edge of the jugular foramen is the opening of the cochlear aqueduct, occupied by the perilymphatic duct, just superolateral to the entry of the glossopharyngeal nerve into the intrajugular part of the jugular foramen. The superior border of the petrous bone or the petrous ridge, grooved by the superior petrosal sinus (connecting the cavernous sinus and the transverse-sigmoid junction), serves as the attachment to the tentorium cerebelli.
The inferior surface of the petrous temporal bone is rough. The apex is connected to the clivus medially by a fibrocartilage and gives attachment to the levator veli palatini and the cartilaginous portion of the eustachian tube. Just posterior to this there is an opening of the carotid canal, which is bounded posteriorly by the carotid ridge (vide supra). The carotid ridge separates the carotid canal from the jugular foramen (Fig. 2.9B).
The petrous temporal bone contains the carotid canal, bony labyrinth, and tympanic cavity (Fig. 2.10B,C). The mastoid air cells are contained in the mastoid part of the temporal bone. The bony labyrinth has three parts: the vestibule, the semicircular canals, and the cochlea. The vestibule, a small cavity in the center of the bony labyrinth, is located at the confluence of the ampullae and the nonampullated ends of the semicircular canals. Medial to it is the meatal fundus, lateral to it is the tympanic cavity, and anterior to it is the cochlea, and it sits on the bony plate harboring the apex of the jugular bulb. The bony plate separating the vestibule and the apex of the jugular bulb is thicker on the left side (average thickness: 8 mm) than on the right side (average thickness: 6 mm).3 This thickness is important during the translabyrinthine approach as the height of the jugular bulb is a major factor determining the degree of exposure. Posterosuperior to the vestibule are the semicircular canals (SCCs). The horizontal or lateral semicircular canal is situated above the tympanic segment of the facial nerve and serves as an excellent guide to locating the tympanic segment of the facial nerve. Henle’s spine, located along the posterosuperior edge of the bony external auditory meatus and anteroinferior to the suprameatal triangle, serves as a good landmark for the lateral semicircular canal, which is approximately 1.5 cm deep to this landmark. The canal is deep to the suprameatal triangle. The posterior semicircular canal lies parallel and close to the posterior surface of the petrous bone just behind and lateral to the lateral end of the internal auditory meatus. This anatomy is important in the retrosigmoid transmeatal approach to vestibular schwannoma. While drilling bone of the posterior edge of the internal auditory meatus to expose the internal auditory canal, the surgeon must be careful not to damage the posterior semicircular canal if the patient’s hearing is to be preserved. The superior semicircular canal projects toward the floor of the middle cranial fossa just below the arcuate eminence. Each of these canals has an ampullated and nonampullated end opening into the vestibule. The ampullae are located at the anterior end of the lateral and superior canals and the inferior end of the posterior canal. The posterior ends of the posterior and superior canals join to form the common crus, which opens into the vestibule. The ampullae of the superior and lateral canals are innervated by the superior vestibular nerve. The posterior ampulla of the posterior canal is innervated by a singular branch of the inferior vestibular nerve. The internal auditory canal (IAC) is medial to the arcuate eminence at an angle of approximately 60 degrees medial from the long axis of the superior semicircular canal.
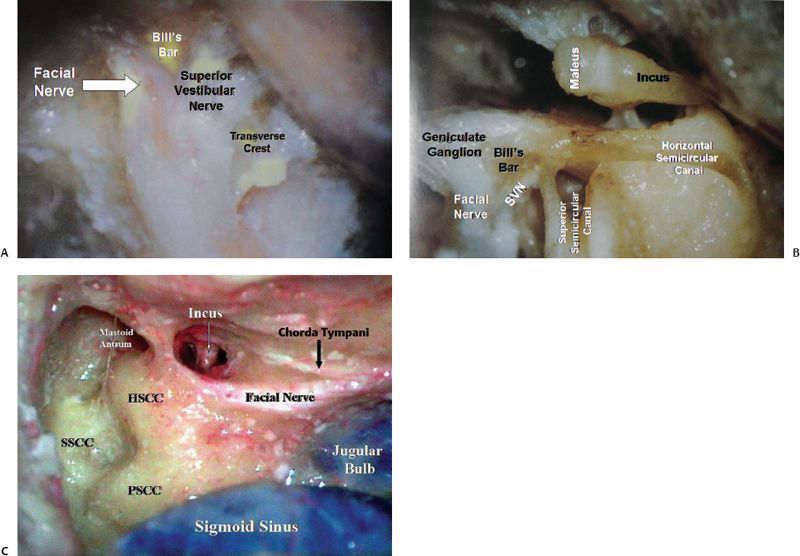
Fig. 2.10 Anatomy of labyrinth. (A) Relation of the nerves in the lateral end of the internal auditory canal. (B) Relationship of the middle ear and labyrinthine structures. (C) Semicircular canals after a total mastoidectomy. HSCC, horizontal semicircular canal; PSCC, posterior semicircular canal; SSCC, superior semicircular canal; SVN, superior vestibular nerve.
The internal carotid artery (ICA) enters the carotid canal and runs vertically upward. The petrous carotid artery starts at the point of entry of the ICA through the periosteal-lined carotid canal in the petrous bone. The external orifice of the carotid canal is directly anterior to the jugular foramen, and its internal orifice is located at the petrous apex. Except at the entrance of the artery to the vertical canal, where it is anchored to the bone by dense bands, the artery can be easily separated from the connective tissue adhesion. The petrous ICA has two segments, the vertical and the horizontal, and they join at the genu. The vertical segment, as the name implies, passes vertically upward in the carotid canal. It is surrounded by the jugular fossa posteriorly, the eustachian tube (ET) anteriorly, and the tympanic bone anterolaterally. The vertical segment turns anteromedially at the genu to form the horizontal segment, which continues anteromedially. It runs anterior to the cochlea, from which it is separated by a thin plate of bone. The anteromedial part of the roof of the horizontal part is formed by the dura or a thin plate of bone, which separates the ICA from the trigeminal/gasserian ganglion. There are one to two branches of the ICA in the carotid canal, and there are a few variations. (Interested readers can find further information in the literature.) A periarterial venous plexus, which is an extension of the cavernous sinus, extends around the petrous ICA for a variable distance. In the majority of the cases, it runs along the anterior and inferior side of the artery, and its extension is limited to the horizontal segment in the majority of the cases. This venous plexus lies within the periosteal covering of the canal, and if the periosteum is intact, bleeding can be avoided. Moreover, the ICA is also accompanied by the sympathetic fibers. The ET and the tensor tympani (TT) lie anterior and parallel to the horizontal segment of the petrous ICA, below the floor of the middle cranial fossa (MCF). Usually the TT lies superior to the ET, and, during dissection, exposure of the TT heralds the imminent exposure of the ET. In most patients, a thin plate of bone separates them. But in many patients they may be separated by fibrous tissue only. In turn, the TT muscle is separated from the carotid canal by a plate of bone, which varies in thickness. Sometimes a part, the superior surface of the muscle, is exposed through a bony dehiscence between the carotid canal and the foramen spinosum (FS). The ET usually is separated from the ICA by a thin layer of bone. The ET crosses the anterolateral aspect of the genu as it exits the middle ear cavity.
The facial nerve in the temporal bone (Fig. 2.10) is divided into four segments: (1) the canalicular segment, located in the IAC, extends from its entry through the IAM up to the meatal fundus; (2) the labyrinthine segment, extending from the meatal fundus to the geniculate ganglion, from where the greater superficial petrosal nerve arises, is situated between the cochlea anteromedially and the semicircular canal posterolaterally; (3) the tympanic segment, running posterolaterally along the medial surface of the tympanic cavity, is situated between the lateral semicircular canal above and the oval window below; and (4) the mastoid segment, which runs vertically from below the midpoint of the lateral semicircular canal, ends by exiting through the stylomastoid foramen. The facial nerve has a very complex course in the temporal bone. The parts of the facial nerve in the temporal bone are part of the IAC, the labyrinthine part, the tympanic part, and the mastoid part. We focus on the labyrinthine part. The canal of Fallopius or the facial canal through the base of the skull is quite long; its course is Z-shaped, threading its way between the labyrinth and the tympanic cavity. This is further complicated by the fact that the segments do not exist in a single plane. The labyrinthine portion begins at the fundus of the IAC, bending forward by approximately 50 degrees the general direction of the IAC. Thus the direction of the canal here is at a right angle to the long axis of the petrous pyramid. It curves forward and inward, skirting the superolateral flank of the basal turn of the cochlea and moving in the direction of the greater superficial petrosal nerve (GSPN). It has a slightly ascending course. Posterolaterally, it comes in close relationship to the superior SCC, where it is sandwiched between the superior SCC and the cochlea, and it is liable to be damaged during drilling of the bone here. The facial nerve now makes a sharp bend posterolaterally, creating the first genu. This is where the geniculate ganglion lies and from here the GSPN arises.
The mastoid part is situated posteriorly in the temporal bone. It projects downward to form the process that is the site of attachment of the sternocleidomastoid, splenius capitis, and longissimus capitis muscles. The lower part medial to the mastoid process is grooved by the digastric notch, to which the posterior belly of the digastric attaches. The posterior border of the mastoid process is perforated by one or more foramina through which an emissary vein to the sigmoid sinus and a dural branch of the occipital artery pass. Posterior to the mastoid part is the asterion, which is the confluence of the lambdoid, occipitomastoid, and parietomastoid sutures. Deep to this is the lower margin of the junction of the transverse and sigmoid sinuses. The upper edge of the junction is just posterior to the junction of the supramastoid crest and squamosal suture. The medial surface of the mastoid part is grooved by the sigmoid sinus, which represents the posterior boundary of the mastoid cavity. At the level of the petrous ridge the sinus meets the roof of the mastoid cavity. The sinodural angle, formed by the angle between the superior petrosal and sigmoid sinus and the middle fossa dura, is an important landmark when performing a mastoidectomy (Fig. 2.10C). The sigmoid sinus courses downward, medially and forward, crossing the occipital bone to enter the jugular foramen.
The medial boundary of the mastoid cavity is formed by the otic capsule, which is a block of bone containing the bony labyrinth. The area of the posterior fossa dura that is exposed through the mastoid cavity between the sigmoid sinus, petrosal sinus, otic capsule, and middle fossa dura is called Trautmann’s triangle. The size of this triangle determines the amount of exposure in the presigmoid retrolabyrinthine approach. The average distance between the anterior margin of the sigmoid sinus and the otic capsule at the level of the posterior semicircular canal in a cadaveric study was 8 mm on the right side and 7 mm on the left side.3 Apart from this distance, the distance between the apex of the jugular bulb and the superior petrosal sinus is also an important determinant in this surgical exposure. The exposure is reduced if the jugular bulb is high. The apex of the jugular bulb is usually inferior to the ampulla of the posterior semicircular canal. The average distance from the apex of the jugular bulb to the superior petrosal sinus in a cadaveric study is 14 mm on the right side and 16 mm on the left side.3 The inside of the mastoid bone is full of air cells, which coalesce to form a cavity called the mastoid antrum, which communicates to the epitympanic part of the tympanic cavity through the aditus. The air cells of the mastoid may extend back behind the sigmoid sinus, up into the squamous part of the temporal bone, anteriorly to the root of the zygomatic process and the floor of the tympanic cavity. The mastoid antrum lies approximately 1.5 cm deep to the suprameatal triangle. The lateral semicircular canal is medial to the epitympanic recess. The posterior semicircular canal faces the medial wall of the antrum. The roof of the antrum is formed by the tegmen in the floor of the middle fossa. The mastoid segment of the facial nerve courses adjacent to the anteroinferior margin of the antrum. The facial nerve has three important sources of blood supply: the AICA provides branches to the proximal segment; the petrosal branch of the middle meningeal artery travels along with the GSPN and supplies the genu, labyrinthine segment, and tympanic segment; and the stylomastoid branch of the posterior auricular artery supplies the mastoid segment. There is good anastomosis between these supplies, and preservation of any two will result in very minimal dysfunction.4
The middle ear cavity or the tympanic cavity (Fig. 2.10B) is a narrow air-filled space between the tympanic membrane and the inner ear. Anteriorly it communicates with the nasopharynx by the ET, and posteriorly it communicates with the mastoid antrum by the aditus. The roof is formed by the tegmen, which separates the tympanic cavity from the middle fossa. The floor separates the cavity from the jugular bulb. The lateral wall is formed by the tympanic membrane and the bony ring to which it attaches. The ring is deficient above, near the openings of the anterior and posterior canaliculi of the chorda tympani nerve. The cavity contains the malleus, incus, and stapes. The tympanic cavity opens up into the epitympanic recess containing the heads of the malleus and incus. The medial aspect of the neck of the malleus is crossed by the chorda tympani nerve. The medial wall has the promontory, the oval window, the round window, and the prominence of the facial nerve. The promontory, which is grooved by the tympanic plexus, represents the basal turn of the cochlea. The apex of the cochlea is medial to the tympanic cavity anterior to the promontory. Posterosuperior to the promontory is the oval window, which connects the tympanic cavity with the vestibule and is occupied by the footplate of the stapes. Above the oval window is the facial nerve prominence. Posteroinferior to the oval window and just below the overhanging edge of promontory is the round window.
♦ Neural Structures
The parts of the brain parenchyma occupying the posterior cranial fossa are the midbrain, pons, and medulla, which constitute the brainstem and the cerebellum (Fig. 2.11).
Midbrain
The midbrain is the uppermost part of the brainstem and it traverses the tentorial hiatus. It is divided in the midline into a right half and left half. Anteriorly and posteriorly deep fissures can be appreciated dividing the midbrain into two halves. Anteriorly the fissure is quite deep to form the interpeduncular fossa. Each right and left half is called the cerebral peduncle. From anterior to posterior each peduncle is subdivided into the crus cerebri or basis pedunculi, the tegmentum, and the tectum. The division between the crus cerebri and the tegmentum is demarcated by the substantia nigra in the parenchyma and the lateral mesencephalic sulcus on the surface. The lateral mesencephalic sulcus is an important intraoperative landmark and is recognized by a longitudinal vein, which runs on the sulcus and connects the basal vein of Rosenthal superiorly and the brachial vein inferiorly. The tectum is behind the aqueduct of Sylvius. Superior and inferior colliculi (quadrigeminal plate) are parts of the tectum and are located on the dorsal aspect of the midbrain. The part superior to the superior colliculus can be a route for entry into the cisterna velum interpositum. The oculomotor nerve exits the midbrain medial to the crus cerebri into the interpeduncular fossa. The plane between the crus cerebri and the oculomotor nerve is a potential place of surgical entry into the midbrain.5 The pontomesencephalic sulcus, separating the midbrain from the pons, starts in the depth of the interpeduncular fossa and runs around the inferior margin of the crus cerebri to join the lateral mesencephalic sulcus. The midbrain continues superiorly with the diencephalic structures. Just above the quadrigeminal plate, at the midline, is the pineal gland. Internally at the third ventricular level, structures inside the posterior third ventricle (posterior commissure, pineal gland, and suprapineal recess) are located above the superior colliculi. The velum interpositum, the space located between the superior and inferior membranes of the tela choroidea, is located just above the pineal gland. The internal cerebral vein and vein of Galen are located just above the pineal gland, and the terminal portion of the vein of Rosenthal is superolateral to the pineal gland.
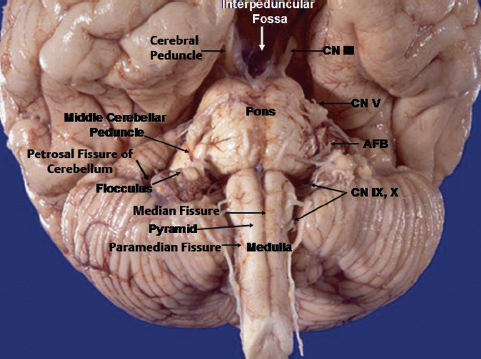
Fig. 2.11 Anterior surface of the brainstem and the petrosal surface of the cerebellum. AFB, acousticofacial bundle; N III, oculomotor nerve; CN V, trigeminal nerve; CN IX, X, glossopharyngeal and vagus nerves.
Pons
The pons is below the midbrain and has a prominent anterior surface, which is convex from side to side. Laterally it is continued in the middle cerebellar peduncle on both sides, which consists of transverse crossed fibers from the pons and higher up. In the midline anteriorly there is a sulcus called the basilar sulcus, which lodges the basilar artery trunk. The lateral pontine sulcus is a shallow groove separating the pontine belly and middle cerebellar peduncle. Just lateral to the lateral pontine sulcus emerges the trigeminal nerve. The trigeminal nerve has two roots. The smaller motor root is in the superomedial position at the emergence, and the larger sensory root is inferolateral in position. For practical microneurosurgical purposes, the emergence of the trigeminal nerve is considered a demarcation point between the pons and the middle cerebellar peduncle. In its posterior part the pons forms the upper part of the floor of the fourth ventricle.
Medulla
The medulla is located inferior to the pons and is separated from the pons by the pontomedullary sulcus. On its anterior part are one median and two paramedian fissures. The median fissure is also called the anterior median fissure, which continues inferiorly as the anterior median fissure of the spinal cord. Just lateral to the anterior median fissure there is a pyramid, which is condensation of the corticospinal tract, on each side. The paramedian fissure, also called the anterolateral sulcus or the preolivary sulcus, is continued below as the anterolateral sulcus of the spinal cord. The rootlets of the hypoglossal nerve come out from the preolivary sulcus. At the upper end of the preolivary sulcus, at its junction with the pontomedullary sulcus, the abducens nerve emerges. Lateral to the preolivary sulcus are the olives on each side, behind which the rootlets of the accessory, vagus, and glossopharyngeal nerves exit from the postolivary sulcus. These nerves extend laterally and exit the cranial cavity via the jugular foramen. Below, the postolivary sulcus is continued as the posterolateral sulcus of the spinal cord. At the upper end of the post-olivary sulcus, there is a triangular depression behind and above the olive and anteromedial to the flocculus representing the junction of the pons, medulla, and middle and inferior cerebellar peduncles. This area, known as the supraolivary fossette, resembles a right triangle. The superior side corresponds to the inferior margin of the pons and middle cerebellar peduncle, the vertical side corresponds to the posterior border of the olive, and the hypotenuse corresponds to the inferior cerebellar peduncle. Cranial nerves VII and VIII exit from the brainstem at the superior end of the supraolivary fossette and then pass above the flocculus to the internal auditory meatus. Cranial nerves IX and X exit from the brainstem at the hypotenuse of the supraolivary fossette and then pass below the flocculus to the jugular foramen.
Cerebellum
The cerebellum wraps the brainstem from the posterior and posterolateral aspects (Fig. 2.9). The fourth ventricle, described as a tent-shaped, midline structure, comes in close contact with the vermian components of the cerebellum. The anatomy of the cerebellum and fourth ventricle are discussed together.
The cerebellum has three surfaces: (1) the petrosal surface, related to the petrous part of the temporal bone anterolaterally; (2) the tentorial surface, related to the tentorium cerebelli superiorly and the upper part of the roof of the fourth ventricle inferiorly; and (3) the suboccipital surface, related to the squamous part of the occipital bone inferiorly and the inferior part of the roof of the fourth ventricle anteriorly.
The suboccipital surface of the cerebellum is located below the lateral sinus and faces inferiorly. In the midline there is the posterior cerebellar incisura and just lateral to that, on two sides, are the velohemispheric or paravermian fissures separating the inferior vermis and cerebellar hemispheres. The parts of the inferior vermis and the corresponding parts of the cerebellar vermis are listed in Table 2.1. The great horizontal fissure starts in the midline between the folium and tuber, and runs laterally between the superior and inferior semilunar lobule. Traced laterally it continues onto the petrosal surface as the petrosal fissure. The secondary fissure is located between the tonsil and biventral lobule. The tonsils are two kidney-shaped structures and are hemispheric components of the uvula (Table 2.1). They are attached to the cerebellum through the superolaterally located peduncles of the tonsil (Fig. 2.12). The tonsil is a relatively free structure, and the superior, anterior, medial, posterior, and most of its lateral surfaces can be separated easily from the surrounding structures. Between its superior pole and the inferior medullary velum is the supratonsillar space; in between the two tonsils is the vallecula; between the anterior surface of the tonsil and the medullar surface is the cerebellomedullary fissure. The cerebellomedullary fissure is one of the routes into the fourth ventricle, instead of splitting the vermis.
| Part of Vermis | Corresponding Part of Cerebellar Hemisphere | |
|---|---|---|
| Superior vermis | Lingula | No hemispheric component |
| Central lobule | Wings of central lobule | |
| Culmen | Quadrangular lobule | |
| Declive | Simple lobule | |
| Inferior vermis | Folium | Superior semilunar lobule |
| Tuber | Inferior semilunar lobule | |
| Pyramid | Biventral lobule | |
| Uvula | Tonsil | |
| Nodule | Flocculus |
The floor of the fourth ventricle (Fig. 2.13) is rhomboidshaped and presents a strip between the lower margin of the cerebellar peduncles and the site of attachment of the tela choroidea. This strip, called the junctional part, is characterized by the medullary striae that extend into the lateral recesses. The junctional part divides the floor of the fourth ventricle into two unequal triangular parts. The larger superior part, with the apex directed toward the aqueduct, is the pontine part, and the smaller inferior part, with the apex directed toward the obex, is the medullary part. The floor is divided longitudinally into two equal halves by the median sulcus. Each half is again divided into two parts by the sulcus limitans or limiting sulcus, which is discontinuous at the junctional area. The medial part is the median eminence and the lateral part is the vestibular area. The sulcus limitans demarcates the motor and sensory areas. The motor nuclei of the cranial nerves are located medial to the sulcus limitans, and the sensory nuclei are located lateral to the sulcus limitans. In the pontine part on the median eminence on each side, just lateral to the median sulcus, there are facial colliculi, which are rounded prominences. Each facial colliculus is bounded laterally by the superior fovea, a dimple formed by the sulcus limitans. The inferior part is characterized by three triangular areas overlying the hypoglossal and vagus nuclei and the area postrema. The areas overlying the hypoglossal and vagal nuclei are the hypoglossal and vagal trigones. Just lateral to the hypoglossal trigone, there is another dimple formed by the sulcus limitans, called the inferior fovea.
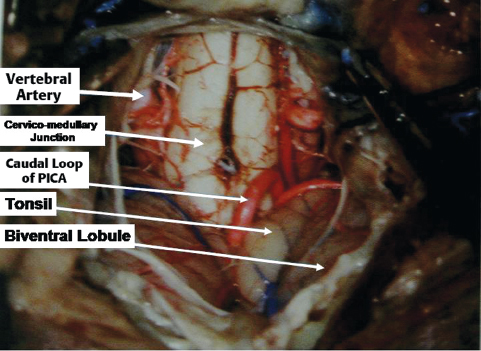
Fig. 2.12 Exposure of the suboccipital surface of the cerebellum during surgery. PICA, posterior inferior cerebellar artery.
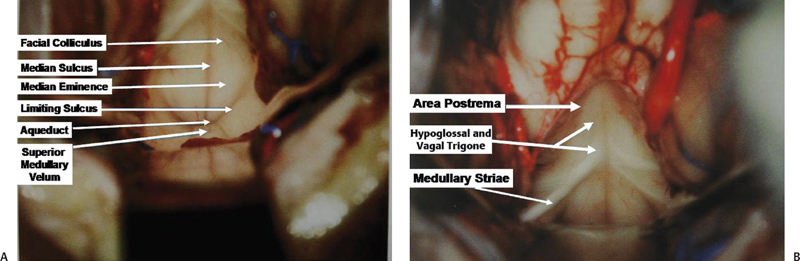
Fig. 2.13 Exposure of the floor of the fourth ventricle during surgery. (A) The upper part of the floor. (B) The lower part of the floor.
The tentorial surface faces the tentorium. It has two incisura. The brainstem fits into the anterior cerebellar incisura and the falx cerebelli fits into the posterior cerebellar incisura. The anterosuperior margin, extending from the top of the culmen downward, forward, and laterally to a point above and behind the middle cerebellar peduncle, forms the posterior wall of the cerebellomesencephalic fissure. The anterolateral and posterolateral margins separate this surface from the petrosal and suboccipital surfaces, respectively. The lateral angle, which is seen in the lateral projection of angiography at the junction of the transverse and sigmoid sinus, is the confluence of the anterolateral and posterolateral margins. The anterior angle is the confluence of the anterosuperior and anterolateral margins. The folia of the tentorial surface are represented by the superior vermis and its hemispheric counterpart (Table 2.1). Also part of the folium and superior semilunar lobule comes into the tentorial surface. The semilunar lobule occupies all the three surfaces of the cerebellum. Most of the cerebellar lobules (except the lingual, tonsil, and nodule) occupy more than one surface. The cerebellomesencephalic or precentral cerebellar fissure is located between the cerebellum and midbrain. It is bounded posteriorly by the culmen above and the central lobule below. The lateral boundary is formed by the anterior surface of the quadrangular lobule above and the wing of the central lobule below. The anterior boundary in the midline is lingual and formed laterally by the superior and middle cerebellar peduncles. Of the cerebellar nuclei (fastigial, globose, emboliform, and dentate), the dentate nucleus is the most laterally located and the largest one. Most of the fibers of the superior cerebellar peduncle originate from the dentate nucleus, which is located at the posterior projection of the superior cerebellar peduncle. The lateral boundary of the dentate nucleus extends 0.5 to 2.0 cm from the midline.6 The lateral boundary of the dentate nucleus roughly correlates well with the posterior continuation of the interpeduncular sulcus (the sulcus between the superior and middle peduncles). This information is important when part of the cerebellum needs to be resected. The deep nuclei of the cerebellum must be preserved at all costs.
The petrosal surface faces the posterior surface of the petrous temporal bone and has two halves because of the interposition of brainstem. The two halves are separated by the petrosal fissure (Fig. 2.11), which runs lateral to medial. At the level of the flocculus, the petrosal fissure bifurcates into a superior limb and an inferior limb. The infrafloccular portion or the inferior limb separates the flocculonodule lobule from the rest of the cerebellum. It communicates with the cerebellomedullary fissure. The folia constituting the upper half of the petrosal surface are those of the tentorial surface that have folded over the middle cerebellar peduncle and the core of the cerebellum; similarly, the folia of the inferior petrosal surface of the cerebellum are formed by folia of the suboccipital surface of the cerebellum, which have folded over the inferior cerebellar peduncle. The choroids plexus and the rhomboid lip of the foramen of Luschka are located anteriorly and inferiorly to the flocculus. Just in front of the choroid plexus is the single superior rootlet of the glossopharyngeal nerve. The flocculus is located immediately below the lateral extension of the pontomedullary sulcus.
The fourth ventricle is a rhomboid-shaped structure, whose overall shape resembles that of a tent that has been turned over, with its base facing anteriorly with its two open lateral walls. The posterior surface of the pons and medulla forms the floor. The superior part of the roof is formed by the superior cerebellar peduncles, the superior medullary velum, and the adjacent lingula. The inferior part of the roof is formed by the inferior medullary velum, tela choroidea, choroids plexus, uvula, and nodule. The two open lateral walls of the fourth ventricle are represented by the lateral recesses through which the fourth ventricle communicates with the cerebellopontine cistern. The conduit is also known as the foramen of Luschka through which the choroid plexus protrudes. The upper half of the roof of the fourth ventricle is composed of neural elements such as the superior cerebellar peduncles, superior medullary velum, and lingula. The lingula is easily visualized through the transparency of the superior medullary velum. The lower half of the roof is composed of nonneural elements and has a horizontal component and a vertical component. The horizontal part is the inferior medullary velum covering the nodule and superior pole of the tonsils. The vertical portion consists of the tela choroidea and the choroid plexus, covering the anterior aspect of the nodule, uvula, and part of the tonsils. These two parts unite at the telovelar junction and continue laterally as the floor of the lateral recess. This telovelar junction is important in gaining entry into the fourth ventricle. Entry through this route precludes splitting of the vermis, which results in severe truncal ataxia. Through the lateral recess the fourth ventricular cavity communicates with the cerebellopontine cistern. The direction of the lateral recess anteroinferiorly and laterally makes an angle of 45 degrees with the sagittal plane. The lateral recess has an anterior wall, superior wall (both formed by the inferior cerebellar peduncle), posterior wall, and floor (formed by the tela choroidea, choroid plexus, and inferior medullary velum from anterior to posterior). The inferior medullary velum becomes thick at the foramen of Luschka where it forms the peduncle of flocculus, which forms the posterior wall of the foramen of Luschka. The choroid plexus of the fourth ventricle is T-shaped with two vertical bars. The horizontal part starts from the fourth ventricle and protrudes into the cerebellopontine cistern. The vertical and proximal parts of the horizontal limb are supplied by the posterior inferior cerebellar artery (PICA), and the rest of the horizontal limb is supplied by the AICA.

Fig. 2.14 The right vertebral artery in the suboccipital triangle is exposed in a cadaver. (A) The muscles of the suboccipital triangle. (B) After removal of muscles. VA, vertebral artery.
♦ Vascular Anatomy
Arteries of the Posterior Fossa
The arteries of the posterior cranial fossa mainly come from the vertebrobasilar system. Arising from the first part of subclavian artery, the vertebral artery enters the foramen transversarium of the C6 vertebra. It then courses up through the transverse foramina of the cervical vertebrae to C2. After exiting from the foramen transversarium of C2, it curves laterally to enter the laterally placed foramen transversarium of C1. The C2 nerve root crosses this segment of the vertebral artery from behind. After exiting from the C1 transverse foramen, the vertebral artery sweeps behind the lateral mass of the atlas, just above the posterior arch in the suboccipital triangle to course superomedially to pierce the dura at the foramen magnum. The posterior spinal artery and posterior meningeal artery emerge from the vertebral artery at this level. At this level, the vertebral artery is accompanied by the suboccipital nerve or C1 nerve.
The vertebral artery can be divided into four parts: the first part extends from the origin to the entry into the foramen transversarium of the C6 vertebra. The second part is inside the transverse foramen of the cervical vertebrae. The third part is in the suboccipital triangle (Fig. 2.14). The fourth part is intradural and can be divided into lateral and anterior medullary segments, before it joins the mate from the opposite side to form the basilar artery. Usually one of the vertebral arteries is the dominant one; usually it is the left one, as it is wider than the right one. From the entrance into the intradural compartment the vertebral artery courses anteromedially and superiorly through the lower cranial nerve rootlets and lateral to the medulla to reach the preolivary sulcus. This segment is the lateral medullary segment of the vertebral artery. This segment begins at the dural foramen just inferior to the lateral edge of the foramen magnum. The dura in this region forms a funnel-shaped foramen around the first 4- to 6-mm length of the proximal part. The first cervical nerve exits and the posterior spinal artery enters the spinal canal through this dual foramen. These structures are bound by fibrous bands. The initial intradural segment of the vertebral artery passes just superior to the first cervical nerve and just anterior to the posterior spinal artery, the dentate ligament, and the spinal accessory nerve. After the preolivary sulcus, the anterior medullary segment begins, which runs in between the hypoglossal rootlets or anterior to them and crosses the pyramid to reach the pontomedullary sulcus, where it unites with the opposite vertebral artery to form the basilar artery. The branches of vertebral artery are the posterior spinal artery, anterior spinal artery, PICA, and anterior and posterior meningeal arteries. Several perforators arise from the vertebral artery, which penetrate the anterior and lateral surface of the medulla (Fig. 2.15).
The PICA arises from the vertebral artery at the anterolateral aspect of the brainstem (Figs. 2.15 and 2.16) near the inferior olive, and it supplies the medulla, inferior vermis, inferior part of the fourth ventricle, tonsils, and inferior aspect of the cerebellum. It is the largest branch of the vertebral artery. It has the most complex relationship with the cranial nerves of any artery, and it is frequently exposed in approaches directed to the fourth ventricle. After its origin it passes posteriorly around the medulla either caudally or rostrally to the hypoglossal rootlets. On reaching the posterolateral margin of the medulla, it passes rostral to the fila of the glossopharyngeal, vagus, or accessory nerves or in between them. After passing these nerves, it winds around the cerebellar tonsil and courses posterior to the roof of the fourth ventricle. After turning away from the roof of the fourth ventricle, it enters a series of deep fissures among the tonsil, vermis, and hemisphere. On exiting the fissures, the branches are distributed to the tonsil and the inferior surface of the vermis and hemisphere. In one study, 12.5% of PICAs passed between the glossopharyngeal and vagus nerves, 20% between the vagus and accessory nerves, and 65% through the rootlets of the accessory nerve.
The PICA is divided into five segments.7 The anterior medullary segment is short, located anterior to the medulla, and extends from the origin to the level of inferior olive. The lateral medullary segment extends from the inferior olive to the origin of cranial nerves IX, X, and XI and is located lateral to the brainstem. The lateral medullary segment of the PICA often has a lateral loop, which in 20% of cases is pressed against the inferior surfaces of the facial and vestibulocochlear nerves. The lateral medullary segment of the PICA passed superior to the hypoglossal nerve in approximately 20% of cases, through the rootlets of the hypoglossal nerve in 47.5%, and inferior to the hypoglossal nerve in 30%.8 The posterior medullary segment (also known as the tonsillomedullary segment) begins at the level of the origin of the cranial nerves IX, X, and XI and loops below the inferior pole of the cerebellar tonsil and upward along the medial surface of the tonsil toward the inferior medullary velum. This is also known as the caudal loop. The next segment, the supratonsillar segment (also known as the telovelotonsillar segment), runs in the cleft between the tela choroidea and the inferior medullary velum superiorly and the tonsil inferiorly. This begins below the fastigium, where the PICA turns posteriorly over the medial side of the superior pole of the tonsil. This part is called the cranial loop. The junction of the posterior medullary segment and the supratonsillar segment is called the choroidal point. The last segment is the cortical segment. After a short distance distal to the apex of the cranial loop, the PICA continues posteroinferiorly in the retrotonsillar fissure. Here it bifurcates into two terminal branches, the tonsillohemispheric and inferior vermian branches. The tonsillohemispheric branch courses inferiorly near the prepyramidal sulcus and gives off anterior or tonsillar branches and posterior or hemispheric branches, which curve posteroinferiorly around the biventral lobule to the underside of the cerebellar hemisphere. The inferior vermian branch lies on the lower aspect of the inferior vermis and forms a convex loop (the pyramidal loop) around the copula pyramidis (connects the pyramid to the biventral lobule and forms the posterior wall of the retrotonsillar space). The anteriormost point of the curve of the pyramidal loop is called the copular point. Apart from terminal branches, the PICA gives rise to perforating, choroidal, and cortical branches; 5 to 20% of PICAs have an extradural origin.9
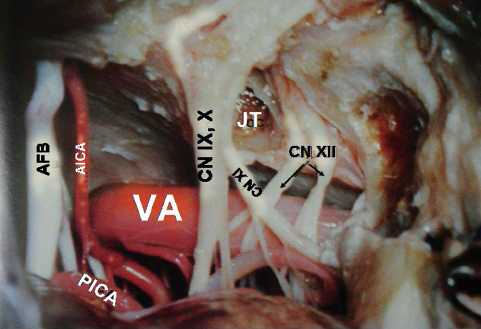
Fig. 2.15 Relationship of the right vertebral artery with the lower cranial nerves. AFB, acousticofacial bundle; AICA, anterior inferior cerebellar artery; CN IX, glossopharyngeal nerve; CN X, vagus nerve; CN XI, accessory nerve; CN XII, hypoglossal nerve; JT, jugular tubercle; PICA, posterior inferior cerebellar artery; VA, vertebral artery.
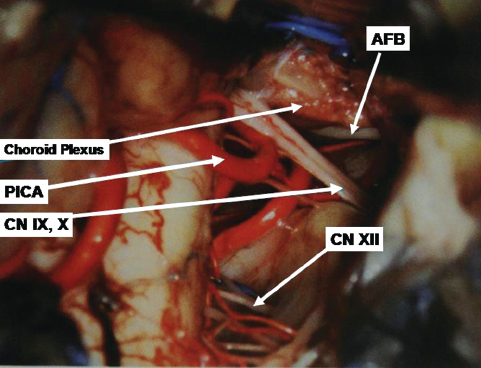
Fig. 2.16 Course of the PICA below the cerebellum.
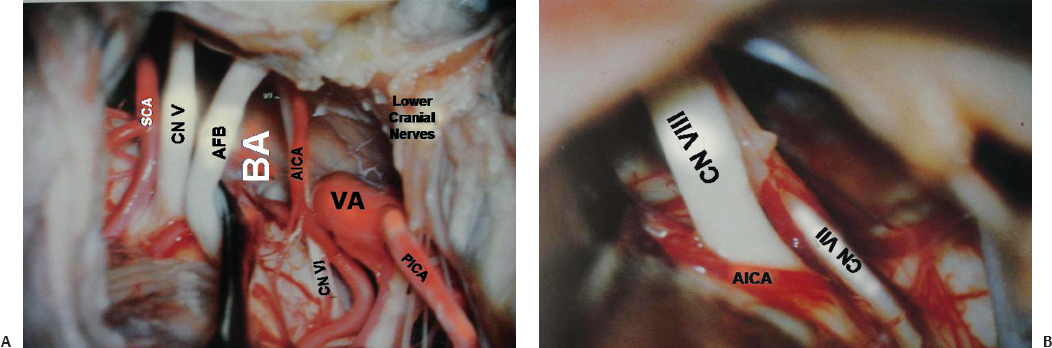
Fig. 2.17 (A) Origin of the AICA and other cranial nerves is shown. (B) Relationship with acousticofacial bundle is shown in more detail. AFB, acousticofacial bundle; AICA, anterior inferior cerebellar artery; BA, basilar artery; CN V, trigeminal nerve; CN VI, abducens nerve; CN VII, facial nerve; CN VIII, vestibulocochlear nerve; SCA, superior cerebellar artery.
The AICA usually arises from lower third of the basilar artery (Fig. 2.17) and less frequently from the middle third. (The AICA and PICA are defined according to their origin rather than by the part of the cerebellum that they supply.) After emerging from the basilar artery, the AICA courses posterolaterally and downward on the belly of the pons in contact with the abducens nerve. It may cross superior or inferior to the abducens nerve. On reaching the acousticofacial bundle, just before or after crossing the rootlets, the AICA bifurcates into its two major branches, the caudomedial and rostrolateral (the main trunk). The caudomedial artery runs inferomedially toward the anterior and medial border of the cerebellum and supplies the middle cerebellar peduncle and biventral lobule. After its origin on the lateral aspect of the pons, the artery runs toward the pontomedullary sulcus to form its caudal loop on the lateral aspect of the pons and medulla. This loop can lie on the anteroinferolateral aspect of the flocculus, on the petrosal aspect of the biventral lobule, or the petrosal aspect of the undersurface of the biventral lobule. Distal to this loop, the next segment (the biventral segment) turns posteroinferiorly to reach the posterior surface of the cerebellum. It may also anastomose with the PICA or give off ascending hemispheric branches, which supplement or may entirely replace the hemispheric branches of the PICA. The rostrolateral trunk runs laterally and curls around the flocculus, and then it courses within the petrosal fissure from where the hemispheric branches to the superior and inferior semilunar lobule are given off.
The rostrolateral trunk is divided into three segments according to their relationship to acousticofacial bundle. The premeatal segment begins at the origin of the rostrolateral trunk and courses around the brainstem to reach the acousticofacial bundle and the region of the meatus. The rostrolateral trunk, with the main trunk of the AICA, forms a caudal loop on the lateral aspect of the pons. The most posteroinferior point is called the caudal point. The rostrolateral trunk then continues superolaterally to reach acousticofacial bundle anterior to the flocculus. On the acousticofacial bundle and the middle cerebellar peduncle it describes either a single arterial or a double arterial loop. The single arterial loop is the single meatal loop because it is related to the internal acoustic meatus. The rostrolateral trunk ascends on the anterior aspect of cranial nerve VII, forming the apex of the meatal loop on the facial nerve near or within the internal auditory canal. From this part occasionally the internal auditory artery is given off. It then reverses the course and descends medially and posteroinferiorly to course between cranial nerves VII and VIII (Fig. 2.17B) or between cranial nerve VIII and the flocculus onto the middle cerebellar peduncle. Here it gives off branches to the middle cerebellar peduncle and the pons. The double arterial loop is called the M segment. The M segment has a proximal meatal loop and a brachial loop closely related to the middle cerebellar peduncle. When the M segment is present, after the meatal loop, the rostrolateral trunk reverses course to ascend as the brachial loop either on the superficial aspect of cranial nerve VIII or on the middle cerebellar peduncle. The apex of the brachial loop is also called the trigeminal point and it can pass anterosuperiorly on the middle cerebellar peduncle to reach the sensory root of the trigeminal nerve. At the trigeminal point it reverses its direction and runs posteroinferiorly on the middle cerebellar peduncle to form the loop. Distal to the single meatal loop or M segment is the postmeatal segment of the rostrolateral trunk. The rostrolateral trunk runs posteriorly and usually inferiorly to enter the suprafloccular portion of the petrosal fissure, where it divides into an ascending branch to the petrosal fissure, which supplies the petrosal surface of the cerebellum, and a descending branch to the posterolateral fissure, which supplies the retrofloccular region.
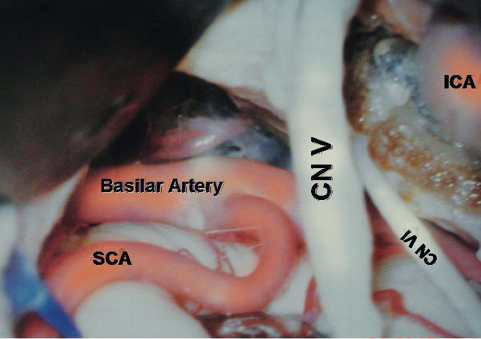
Fig. 2.18 Origin and proximal part of the superior cerebellar artery (SCA). ICA, internal carotid artery.
The superior cerebellar artery (SCA) arises near the apex of the basilar artery (Fig. 2.18) just proximal to the origin of the posterior cerebral artery (PCA) and is the rostral-most artery of the infratentorial compartment. The SCA encircles the lower midbrain and pons and supplies the tentorial surface of the cerebellum, the deep cerebellar nuclei, the upper brainstem, and the inferior colliculi. The SCA can be divided into four segments. From its origin to the anterolateral part of the brainstem is the anterior pontomesencephalic segment. It runs laterally on the anterior aspect of the upper pons, often with a convex arc inferiorly. At the anterolateral margin of the brainstem it lies inferior to the third cranial nerve. The initial course of this segment depends on the site of the basilar bifurcation. With a low bifurcation, this segment passes upward, whereas in high bifurcation, it initially passes anteroinferiorly to reach the anterior surface of the pons. The lateral pontomesencephalic segment starts from the anterolateral margin of the brainstem and runs caudally on the lateral side of the upper pons in the infratentorial portion of the ambient cistern to end at the anterior margin of the cerebellomesencephalic fissure. From this segment arises the first major cortical branch of the SCA, which runs anterolaterally to reach the anterolateral margin of the cerebellum. It then runs posterolaterally in the region of the petrosal fissure to supply the adjacent areas. Its area of supply is inversely related to the area supplied by the AICA. This vessel is an important angiographic landmark to locate the anterolateral margin and anterior angle of the cerebellum. Medial to this segment is the brainstem; lateral to this segment is the wing of the central lobule; inferior to this segment is middle cerebellar peduncle. Its caudal loop projects toward and often reaches the root entry zone of the trigeminal nerve. The bifurcation of the SCA into rostral and caudal branches occurs in this segment. The rostral branch supplies the superior vermis and the inner part of the tentorial surface of the cerebellum. The caudal branch supplies the rest of the tentorial surface not supplied by the rostral branch. The third segment, the cerebellomesencephalic segment, runs in the cerebellomesencephalic fissure and reaches the anterosuperior margin of the cerebellum through a series of hairpin bends. From this segment small cortical branches from the rostral and caudal branches arise, and these are precentral branches (twigs from the rostral branch supply the inferior colliculi and superior medullary velum, and twigs from the caudal branch supply the deep cerebellar nuclei). The last segment is the cortical segment. The hemispheric and vermian branches represent this segment.
Veins of the Posterior Fossa
The veins of the posterior fossa can be divided into three groups: (1) the petrosal group or the anterior group draining into the superior and inferior petrosal sinuses, (2) the superior or the galenic group draining into the vein of Galen, and (3) the posterior or tentorial group draining into the venous sinuses in and around the torcular sinus. Usually the veins tend to drain into the venous sinus closest to them. The exceptions are the veins on the surface of the midbrain, which drains into the galenic system. The veins of the posterior fossa are summarized in Table 2.2.
♦ Cisternal Anatomy
There are several subarachnoid cisterns in the posterior fossa or infratentorial compartment. Understanding the anatomy and disposition of these cisterns is extremely important in performing microsurgical dissection (Figs. 2.15, 2.16, 2.17, 2.18, and 2.19).
The interpeduncular cistern is a cone-shaped cul-de-sac occupying the interpeduncular fossa and is bounded above by the inferior surface of the mesencephalon, the lower diencephalons, the posterior perforated substance, and the mamillary bodies. Anteroinferiorly it is bounded by the clivus and superolaterally it is bounded by the carotid and crural cisterns and the medial temporal lobes. Inferolaterally it is continuous with the cisterna ambiens. The anterosuperior wall of this cistern is thick. Like a curtain, it stretches from one medial temporal surface to another and is fused with the chiasmatic cistern around the infundibulum and pituitary stalk. Above, it is attached to the mamillary bodies, and inferiorly it is attached to dorsum sellae. This is Liliequist membrane. The inferior part of the cistern extends in a triangular shape down to the middle portion of the basilar artery. The origins of the PCA and SCA are in this cistern. Within the interpeduncular cistern, the PCA and SCA do not have much arachnoid membrane around them, but at the level of the oculomotor nerve, the PCA and SCA acquire their own arachnoid sleeve and enter the cisterna ambiens. The contents of the interpeduncular cistern are listed in Table 2.3.
Table 2.2 Veins of Posterior Fossa
| Major Group | Divisions | Veins |
|---|---|---|
Petrosal* group | Veins related to the anterior aspect of the brainstem | • Anterior pontomesencephalic vein |
• Transverse pontine vein | ||
• Lateral pontine vein | ||
• Anterior medullary vein | ||
• Parenchymal perforating vein | ||
Veins in the wing of precentral cerebellar fissure | • Brachial veins | |
Veins on the superior and inferior surfaces of the cerebellar hemisphere | • Superior hemispheric veins, lateral group (draining lateral portion of the wing of central lobule, quadrangular lobule, simple lobule, tentorial part of superior semilunar lobule) | |
• Inferior hemispheric veins | ||
Includes veins of great horizontal fissure | ||
Veins on the cerebellar side and medullary side | • Medial tonsillar vein | |
• Retro-olivary vein | ||
• Vein of inferior cerebellar peduncle | ||
Vein of the lateral recess of the fourth ventricle | ||
Superior or galenic group | Mesencephalic tributaries | • Median anterior pontomesencephalic vein |
• Lateral anterior pontomesencephalic vein | ||
• Lateral pontomesencephalic vein | ||
• Lateral mesencephalic vein | ||
• Peduncular vein | ||
• Posterior mesencephalic veins | ||
• Tectal veins | ||
Cerebellar tributaries | • Precentral cerebellar vein | |
• Superior vermian vein (draining central lobule and culmen) | ||
• Superior hemispheric veins, anterior group (draining intermediate portion of the wing of the central lobule and quadrangular lobule) | ||
Tentorial or posterior group | Inferior vermian vein | • Superior retrotonsillar vein |
• Inferior retrotonsillar vein | ||
Drains tuber, pyramid, uvula | ||
• Superior hemispheric veins, posterior group (draining declive, folium, intermediate portion of simple and superior semilunar lobules) | ||
• Inferior hemispheric veins |
*Superior petrosal vein is formed by the junction of the transverse pontine and brachial veins, and the vein of great horizontal fissure.
The prepontine cistern is located between the anterior surface of the pons and the clivus and surrounds the basilar artery. The AICA originates in this segment, and moderately dense arachnoid fibers encircling the AICA denote its exit from this cistern into the cerebellopontine cistern. Laterally, the cistern is bounded by an arachnoid membrane, which forms the medial boundary of the cerebellopontine cistern. Inferiorly, the arachnoid wall of the cistern is thickened at the confluence of the vertebral arteries forming the basilar artery. Superiorly, a plane separates the prepontine cistern from the interpeduncular cistern. The contents are listed in Table 2.3.
The premedullary cistern extends superiorly from the pontomedullary sulcus to the upper cervical area inferiorly. The cistern’s anterior boundary is the clivus, and the posterior boundary is the anterior surface of the medulla.
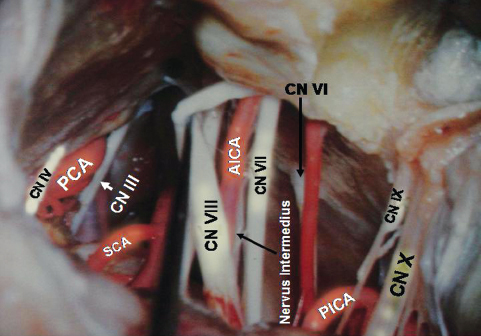
Fig. 2.19 Posterior fossa cranial nerves are on display. CN III, oculomotor nerve; PCA, posterior cerebral artery.
The ambient cistern (cisterna ambiens) skirts the lateral aspect of the mesencephalon and has both supratentorial and infratentorial components. It is a paired structure. Medially it is bounded by the cerebral peduncle and interpeduncular cistern. Laterally it is bounded by the medial temporal lobe (supratentorial) and the quadrangular lobule of the cerebellum (infratentorial). Inferiorly it shares the wall with the cerebellopontine cistern. Anteriorly it is related to the crural cistern. The superior extension of the ambient cistern, although not in the infratentorial compartment, warrants special mention. It is called the wing of the ambient cistern, which includes the portion of the cistern extending from the uncus of the temporal lobe, over the pulvinar of thalamus, and anteromedially to the area of velum interpositum. The contents are listed in Table 2.3.
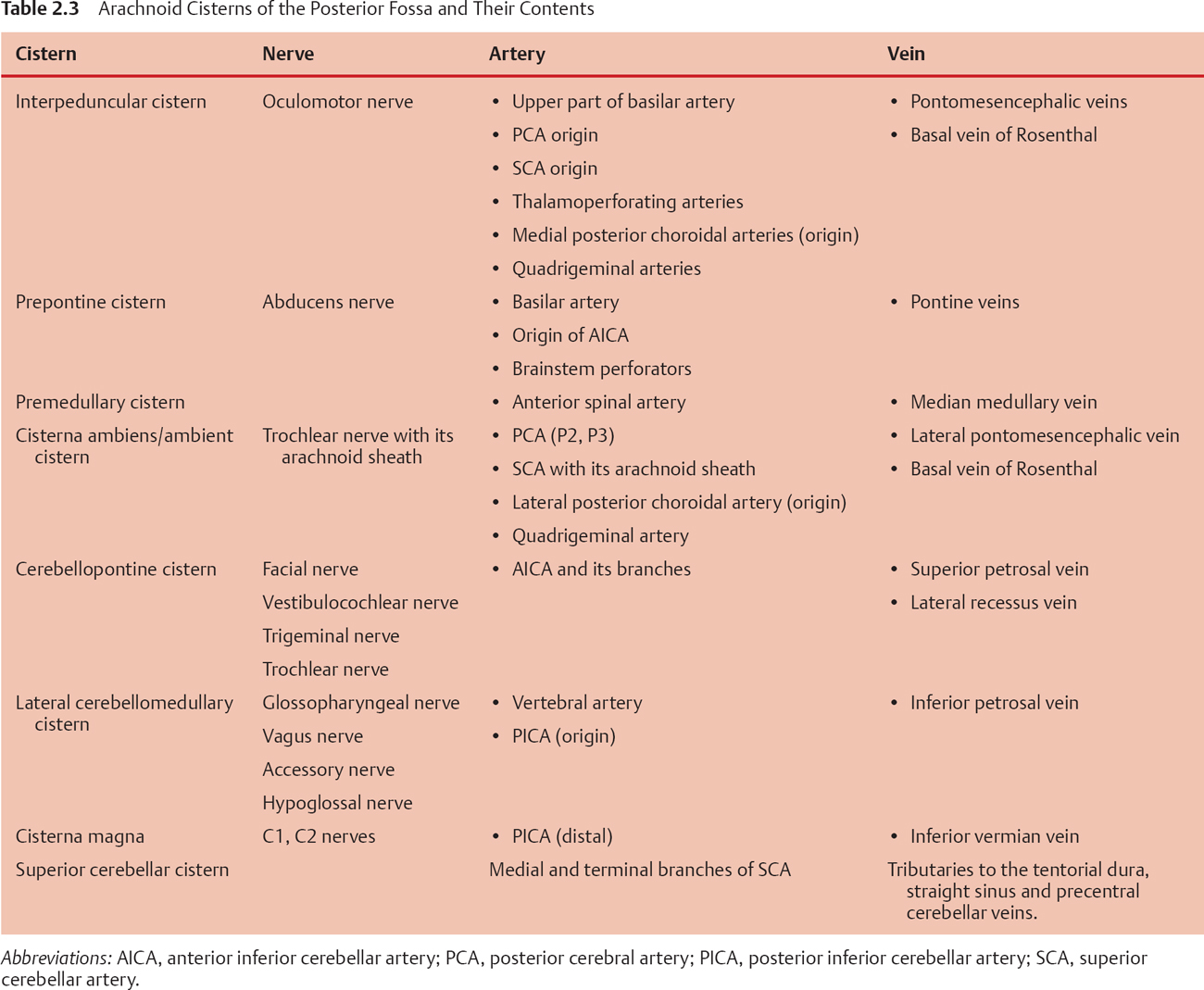
The lateral cerebellomedullary cistern is also a paired structure and is located anterolateral to the medulla. The anterosuperior border is the sulcus between the pons and medulla. Arachnoid over the glossopharyngeal, vagus, and accessory nerves separates this cistern from the cisterna magna dorsally and from the cerebellopontine cistern superiorly. A less clear arachnoid layer separates it from the premedullary cistern. The cistern extends from the pontomedullary sulcus to the foramen magnum. Sleeves of arachnoid run along with the glossopharyngeal, vagus, and accessory nerves to the respective exit foramen.
The cisterna magna can be considered the upper widened extension of the dorsal spinal subarachnoid space. The cistern is bounded anteriorly by the dorsal surface of the upper spinal cord and lower medulla, and extends to the posterior medullary velum. In the midline it runs superiorly beneath the vermis between the tonsils to communicate with the cavity of the fourth ventricle via the foramen of Magendie, with a cephalad extension of the cistern called the vallecula. The cistern has a variable extension dorsally over the vermis. Usually it ends in the region of the lobulus pyramis of the vermis, but it may extend up to the tentorium. Posteriorly the cistern conforms to the inner table of the occipital bone except in the midline, where the falx cerebelli partially divides the cistern. The lateral boundary of the cistern over the cerebellum is the point of fusion of the arachnoid and pia of the cerebellum and that over the brainstem is the arachnoid over the lower cranial nerves forming the posterior boundary of the cerebellomedullary cistern. Tough trabeculae are seen in the cisterna magna stretching between the dorsal medulla and the posterior arachnoidal wall of the cistern. Similar arachnoid fibers stretch between the medulla, cerebellar tonsil, and the ipsilateral PICA. Sometimes a median sheet of arachnoid divides the cistern into sagittal halves, and at the level of C1-C2, two additional paramedian septa are formed and extend to the level of D11-D12. The contents of this cistern are listed in Table 2.3.
The superior cerebellar cistern covers the superior vermis and blends laterally with the subarachnoid space over the cerebellar hemispheres. Anteriorly it extends to the tentorial surface and communicates with the quadrigeminal and ambient cistern.
♦ Anatomy of Surgical Approaches to the Posterior Fossa
Telovelar Approach to the Fourth Ventricle10
A posterior midline incision is made from the lambda to the tip of C2. Suboccipital muscles are dissected and retracted laterally. A suboccipital craniotomy is made. The lateral exposure should be adequate, but exposure of the lateral sinus is not necessary in approaching the fourth ventricle. However, the foramen magnum must be deroofed adequately laterally. Occasionally, removal of the posterior arch of the atlas is needed when the operation is performed with the patient in a semi-sitting position. The dura is opened in a Y-shaped manner. The suboccipital surface of the cerebellum comes into view. The space around the tonsil, like the vallecula, the supratonsillar space, and the cerebellomedullary fissure, is dissected, and the tonsil is freed from the opposite tonsil uvula, inferior medullary velum, tela choroidea, and posterior medullary segment of the PICA. The tonsil is then retracted laterally, which exposes the roof of the fourth ventricle. The tela choroidea or the inferior medullary velum can be resected to gain better access. Occasionally, removal of the tonsil may be necessary.
Supracerebellar Infratentorial Approach to the Posterior Third Ventricle
The space between the tentorium and the tentorial surface of the cerebellum is a wide and excellent natural pathway to access the posterior third ventricular region. The Operation is usually done with the patient in a sitting position to take the advantage of gravity. A midline incision is made from the lambda to the tip of C2. The suboccipital muscles are reflected and retracted laterally. A suboccipital craniotomy is made. It is not always necessary to deroof the foramen magnum, but if the brain is full, the foramen magnum should be deroofed to release CSF from the cisterna magna. A craniotomy must be made exposing the torcular and transverse sinus, so that the dura can be reflected above to improve visualization. The dura is opened in a curved manner based on the lateral sinus and is reflected upward. The upper part of the dura in the craniotomy opening should be opened. Keeping the lower part intact prevents herniation of the cerebellum through the craniotomy opening. This also exposes the upper suboccipital surface of the cerebellum and part of the tentorial surface. The torcular and transverse sinus are further retracted superiorly using sutures. The arachnoidal adhesions between the cerebellum and the torcular and transverse sinus are cut. This results in slackening of the cerebellum, and gravity causes it to sag downward, making the supracerebellar space wide. The small bridging veins in the surgical corridor are coagulated and cut. Depending on the pathology and the need of access, both the midline and lateral corridors may be used. In the midline corridor between the vermis and the straight sinus in the tentorium, the culmen is the main obstacle. The culmen is retracted below. Often the superior vermian vein or the precentral cerebellar vein or both in the cerebellomesencephalic fissure have to be sacrificed. The arachnoid in the quadrigeminal cistern is very thick and must be opened carefully, because there is a chance of injury to the branches of the SCA and PCA and even the great veins. Usually after opening the arachnoid adequately the lesion in the posterior third ventricle comes into view.
Preauricular Subtemporal-Infratemporal Approach to the Petroclival Region11,12
This approach is used mainly to access the extradural petroclival junction, and access can be made from the nasopharynx to the foramen magnum. After the skin and the subcutaneous tissue are mobilized in the temporal region, parotidomasseteric fascia is dissected and separated from the parotid gland. This maneuver is extremely important. Later, when the mandibular condyle is retracted downward, the facial nerve is liable to become stretched. To minimize the traction on the facial nerve, the parotidomasseteric fascia is dissected free. The zygomatic arch and the frontozygomatic process are completely denuded of periosteum and exposed. A temporal or a frontotemporal craniotomy is made. Under the microscope the temporal dura is separated gently from the lateralmost part of the MCF. While separating, the arcuate eminence (AE) is identified. This is usually the posterior boundary of the dissection. The dural separation is then continued anteriorly to reach the foramen spinosum where the middle meningeal artery (MMA) is seen entering the cranial cavity. Further forward dissection exposes the lateral rim of the foramen ovale transmitting the mandibular division of the trigeminal nerve (V3). At this stage it is not possible to see the V3 clearly as it is covered by dural reflections. Medial dissection in between the AE and the MMA exposes the lesser superficial petrosal nerve (LSPN), which can be seen coursing toward the foramen ovale. The LSPN is cut and medial separation of the dura is continued. The MMA is cut at this stage. Further medial dissection exposes the GSPN emerging from the facial hiatus to supply part of the facial nerve. Care must be taken while exposing the GSPN, because in most cases there is no bony roof and the dural fibers are intermingled with the nerve. The nerve must be searched for, and the dura mater is gently separated (often with sharp dissection) from the nerve. It is important to remember that if the GSPN is avulsed while separating the dura mater, the major brunt will be borne by the facial nerve, not the GSPN, which will be cut in any case.
The osteotomy is now made. Anteriorly, a V-shaped cut is made at the level of the frontozygomatic and the zygomaticomaxillary suture. The posterior cut is made to include the condylar fossa. Here also the cut is V-shaped, with the apex of the V falling just short of the foramen spinosum and the limbs spanning the anterior and posterior extent of the zygomatic root. Before making the osteotomy, the capsule of the temporomandibular joint is exposed (which can be facilitated by slight anteroinferior retraction of the masseter muscle) and cut sharply. The meniscus of the joint is separated from the condylar fossa. The posterior cut of the osteotomy is then made. While making the osteotomy, one must be careful not to enter the middle ear cavity. The zygomatic arch is then removed. This exposes the mandibular condyle covered by the meniscus. The dural attachment to the V3 nerve is sharply cut and separated from the V3. This exposes a venous plexus covering the V3. Dissection is continued anteriorly to reach the V2 exiting the foramen rotundum. The venous plexus around these nerves, which are extensions of the cavernous sinus and the venous plexus around Meckel’s cave, is common. The GSPN may be cut at this stage, if desired, close to the facial hiatus. Further dissection of the dura mater medially exposes part of the horizontal segment of the petrous ICA, close to the V3. The horizontal part of the petrous ICA is variably roofed with bone. The part of the MCF lateral to the GSPN, the V3, and the V2 is drilled. This exposes the lateral part of the root of the pterygoid process. Drilling the bone of the MCF between V2 and V3 may expose the sphenoid sinus.
Attention is then turned toward exposure of the petrous ICA. Initially a search is made posteroinferomedial to V3, whether or not any part of the ICA is visible. If it is visible, it serves as a good guide. If the ICA is completely covered by bone, one must take extra caution while drilling. Drilling lateral to the horizontal part of the petrous ICA is relatively safe. The ICA is covered by a venous plexus variably. During drilling, bleeding from this venous plexus may occur. But if the periosteal sheath covering the ICA is kept intact, bleeding does not occur. While drilling the bone laterally, one first sees the muscle and glistening tendon of the tensor tympani muscle (TTM). The muscle is cut and reflected away. Just inferior to the TTM, the cartilaginous part of the ET is found. The ET may be separated from the ICA by a very thin plate of bone. Dissection is made around the ET, to free it from the ICA, and then it is cut to expose the lateral wall of the horizontal part of the petrous ICA completely. Bone drilling is continued to expose the superior, lateral, and inferior walls of the horizontal part of the petrous ICA. During exposure of the genu, care must be taken to avoid damage to the cochlea. However, if the drilling is done lateral to the ICA, the chance of injuring the cochlea is minimal. In this approach, it may not be necessary to drill the bone superior to the genu completely. The cochlea is posteromedial to the genu.
The petrous bone and the tympanic plate lateral and anterior to the vertical limb of the petrous ICA are drilled completely. During this part of the procedure, the condyle of the mandible has to be retracted inferiorly, and the downward tilt of the vertex is reduced. It may be necessary to tilt the vertex a little upward. This improves the view of the lower part of the petrous ICA. Here, a thick fibrocartilaginous band is found, which is continuous with the periosteum covering the ICA. This fibrocartilaginous ring is cut and is gently dissected away from the wall of the ICA. This frees the ICA to be retracted anteriorly and laterally. However, a thin bone plate, between the V3 and the horizontal part of the petrous ICA, will hinder complete anterior mobilization. This bone forms the medial rim of the foramen ovale. This bone is removed. Then the ICA is completely free to be retracted anteriorly. One or two tapes are passed around the ICA, and the ICA is retracted anteriorly and laterally. Part of the petrous bone medial to the carotid canal is left now. This can be drilled freely as hardly any important structure is there. One may expect some bleeding here during drilling. However, while drilling posteriorly, one must be careful not to go too far back and encounter the cochlea. Further medially and backward the anterior wall of the IAC is seen. The petrous pyramid is drilled completely to expose the posterior fossa dura bounded above and below by the superior petrosal sinus (SPS) and below by the inferior petrosal sinus (IPS). Just below the IPS, the clivus is visible. The dura can be opened along the region of Meckel’s cave or according to the demands of the patient’s condition. Usually the dura is opened in the posterior fossa first. Then we can see a triangular space exposed in the posterior fossa dura bounded inferiorly by the IPS and the clivus bone, superiorly by the trigeminal root and the SPS, and posteriorly by the superior SCC, IAC, and the jugular bulb. The dura is opened in a triangular fashion, with the base posteriorly placed. If necessary, the triangle can be split in two by incising the dura along the anterior aspect of the IAC. This exposes cranial nerve VI first exiting the pontomedullary junction on the ventral surface. The contralateral VI nerve can also be visualized, but the origin may not be. The midbasilar artery, the origin of the AICA, the ipsilateral cranial nerve VII-VIII and IX-X-XI complexes and cranial nerve XII can be seen. In fact it is possible to see the cranial nerve XII of both sides. Intradural pathology ventral to brainstem can be removed by this approach. To gain exposure above, the tentorium is cut. First the dura is cut along the inferior surface of the temporal lobe in a line along the anteroposterior axis. Another incision is made at a right angle to the first one, forming a T. The incision parallel to the coronal plane meets the SPS. The SPS is ligated and cut. Now the temporal lobe is gently elevated to expose the tentorial notch. Cranial nerve IV is seen diving below the tentorium. The tentorium is incised posterior to the point at which nerve IV is diving below the tentorium. The tentorial incision is extended to the point where the SPS was ligated. Sutures are placed in the tentorium and the tentorium is retracted. Now all the cranial nerves from III to XII are exposed. Cranial nerve V is seen exiting the brainstem and entering Meckel’s cave, which has already been drilled.
Retrosigmoid Approach to the Cerebellopontine Angle
This approach, one of the major workhorse approaches to the posterior fossa, is directed through an opening located just behind the sigmoid sinus and is guided down the plane between the posterior face of the petrous temporal bone and the petrosal surface of the cerebellum.13 As mentioned before, during a craniotomy the asterion is an important landmark in localizing the genu of the lateral sinus. While operating on vestibular schwannoma through this approach, it is necessary to remove the posterior meatal wall. In that case it is often necessary to sacrifice the subarcuate artery because it passes through the dura on the posterior meatal wall to reach the subarcuate fossa. This artery has enough length so that it can be coagulated safely without jeopardizing AICA, but sometimes the stem is too short or the loop of the AICA along with the subarcuate artery is incorporated into the dura. In that case, a cuff of dura should be cut to separate the dura and the artery from the posterior meatal lip to prepare the opening of the meatus. The posterior semicircular canal (PSCC) and its common crus with the superior semicircular canal are situated just lateral to the posterior meatal lip, and should be preserved while exposing the IAC by drilling the posterior meatal lip, if hearing preservation is an issue. Similarly, injury to the endolymphatic sac, which expands under the dura on the posterior face of the petrous temporal bone inferolateral to the posterior meatal lip, should be avoided; it may be entered while removing the dura from the posterior meatal lip. Injury to the vestibular aqueduct, situated inferolateral to the posterior meatal lip, should also be avoided. An unusually high jugular bulb may block access to the posterior meatal lip. Encountering mastoid air cells is common while drilling this region and they must be obliterated meticulously to prevent CSF leak.
After the removal of the posterior wall of the internal auditory canal, the dura lining the canal is slit open to expose the contents. The nerves in the lateral part of the IAC are the facial, the cochlear, and the superior and inferior vestibular nerves. The position of the nerves is most constant in the lateral portion of the canal, which is divided into a superior and an inferior portion by a horizontal ridge, called the transverse crest or the falciform crest. The facial and the superior vestibular nerve are superior to the crest. The facial nerve is anterior to the superior vestibular nerve here and is separated from the superior vestibular nerve by Bill’s bar (vide supra). The cochlear and the inferior vestibular nerve run inferior to the transverse crest with the cochlear nerve anterior to the inferior vestibular nerve. Knowing this relation is of utmost importance in surgery of vestibular schwannoma. The tumor usually displaces the facial and cochlear nerves anteriorly. Variable growth of the tumor may displace the facial nerve anteriorly, anterosuperiorly, or anteroinferiorly. It is easier to locate the facial nerve at the lateral end of the IAC, as the relation between the four nerves is most constant here, rather than in the medial location. The landmarks that are helpful in guiding the surgeon to the junction of the facial nerve with the brainstem are the pontomedullary sulcus, the junction of cranial nerves IX, X, and XI with the medulla, the foramen of Luschka with its choroid plexus, and the flocculus. The facial nerve arises from the brainstem near the lateral end of the pontomedullary sulcus approximately 1 to 2 mm anterior to the point where the vestibulocochlear nerve joins the brainstem, which is at the lateral end of the pontomedullary sulcus (where it joins the foramen of Luschka). At this point the interval between the facial nerve and the vestibulocochlear nerve is the greatest.13 Another way of locating the medial end of the facial nerve is to project an imaginary line along the brainstem attachment of cranial nerves IX, X, and XI to the brainstem, and the junction of facial nerve is at the junction of the pontomedullary sulcus and this imaginary line, which is approximately 2 to 3 mm rostral to the rostral-most rootlet of the glossopharyngeal nerve. Distal to the junction with the brainstem, the facial nerve and the vestibulocochlear nerves come closer as they approach the IAM. Apart from the facial nerve, the vestibulocochlear nervus intermedius is also a component of the acousticofacial bundle. There are three segments of nervus intermedius: the proximal segment adheres closely to the vestibulocochlear nerve, the intermediate segment lies free between the motor root of the facial and vestibulocochlear nerves, and distal segment joins the motor root to form the facial nerve.14 In most instances, it arises as a single root at the brainstem anterior to the superior vestibular nerve. Even when it arises as multiple rootlets, it quickly converges to a single trunk before joining the motor root of the facial nerve.
Retrolabyrinthine Petrosal Approach to the Cerebellopontine Angle or the Petroclival Region
This presigmoid approach starts with a standard mastoidectomy. The bone removal extends from the supramastoid crest superiorly, the mastoid tip anteriorly, and the posterior wall of the external auditory canal anteriorly. A straight cut made along the temporal line posteriorly into the sinodural angle delineates the upper portion of the dissection. A second cut is made perpendicularly to the first and toward the mastoid tip. This cut is made immediately posterior to the canal wall. The mastoid cortex is then removed in a systematic fashion, with the deepest penetration occurring at the junction of the two perpendicular lines. The mastoid cortex should be unroofed from the posterior canal wall back to and slightly beyond the sigmoid sinus and at an adequate distance into the mastoid tip.15 This results in a bean-shaped cavity, with the inferior portion formed by the mastoid tip below the sigmoid sinus and the external auditory canal, and the upper portion above the sigmoid sinus extending posteriorly into the sinodural angle and anteriorly into the zygomatic root. The sigmoid sinus, the sinodural angle, and the middle fossa dura should be exposed. Exposure of the middle fossa dura is essential to achieve the best possible access into the antrum and the epitympanic areas.
The next step exposes the mastoid antrum, which is invariably present, although its size may vary. It lies immediately below the deepest point of penetration into the temporal bone, posterior to the spine of Henle and the zygomatic root. Occasionally, before the exposure of the antrum, Körner’s septum, which is actually a segment of the petrosquamous suture line, representing the fusion of the squamous and petrous bone (extending from posterior canal wall at the tympanomastoid suture line and blends with the air cells in the immediate proximity of the middle fossa dural plate), is usually observed. The antrum is identified as a large, air-containing space, at the bottom of which lies the basic landmark of the labyrinthine bone, the horizontal or lateral SCC. Just anterior to the lateral SCC is the aditus through which the middle ear can be seen. Here we can also find the facial buttress. Next, the fossa incudis, the epitympanum (anterior and superior to lateral SCC), and the external genu of the facial nerve (medial and inferior to lateral SCC) are exposed. Drilling the inferior mastoid cells will expose the digastric ridge, which is the cortical indentation of the attachment of the digastric muscle. Traced anteriorly, the digastric ridge leads to the facial nerve at the stylomastoid foramen. Thinning of the posterior wall of the external auditory canal exposes the bone in the facial recess area, bounded medially by the vertical mastoid segment of the facial nerve, laterally by the chorda tympani nerve, superiorly by the fossa incudis, and anteriorly by the tympanic membrane. Unless the facial nerve is transposed, it is prudent to preserve a thin shell of bone over the facial nerve, thereby preserving the periosteum. Then the vessels over the facial nerve can be seen. Preservation of these vessels is of the utmost importance if facial nerve function is to be preserved. Drilling the infralabyrinthine cells exposes the jugular bulb. To expose the jugular bulb completely, the facial nerve needs to be transposed anteriorly by mobilizing. The disorders that require the retrolabyrinthine petrosal approach do not require exposure of the jugular bulb.
Drilling the bone posterior to the lateral SCC exposes the posterior SCC, which is bisected by the lateral SCC, and runs parallel to the posterior fossa plate. The posterior SCC starts from just medial to the facial nerve below the external genu and ends superiorly at the common crus with the superior SCC. The endolymphatic sac, which is medial to the sigmoid sinus and inferior to the posterior SCC, is found by thinning the posterior fossa dural plate, immediately medial to the sinus. It is identified by the presence of thickened white dura next to the darker, single-layered dura. The superior SCC is oriented perpendicularly to the lateral SCC and is approximately parallel to the floor, with its long axis orthogonal to the long axis of the petrous bone. It originates deep to the lateral SCC and arches posteriorly and superiorly to join the common crus. Next, a temporal craniotomy is added with its posterior boundary extending beyond the transverse sinus and the sigmoid sinus junction. The posterior fossa dura is opened first and the CSF in the cerebellopontine cistern is released to make the brain slack. A longitudinal incision is made in the inferior temporal dura. Here the utmost care is needed to preserve the vein of Labbé. The dural incision is then extended in a T-shaped manner to the superior petrosal sinus, which is ligated and the incision is extended medially to cut the tentorium up to its edge, with a special care to preserve the trochlear nerve. This exposes the cerebellopontine angle from the anterolateral direction, and it makes the approach to the clivus shallow. To increase or widen the exposure, the approach can be enlarged by converting it to a partial labyrinthectomy petrous apicectomy approach, a translabyrinthine approach, or a total petrosectomy approach.
Partial Labyrinthectomy Petrous Apicectomy Approach to the Cerebellopontine Angle or the Petroclival Region15,16
The bone removal initially is exactly like that in the retrolabyrinthine approach. After completing the temporal craniotomy, the middle fossa floor is dissected to locate the GSPN, which acts as a guide to the position of the petrous ICA while drilling the medial petrous bone. The superior SCC and part of the posterior SCC are removed by drilling. Medial petrous bone is drilled up to the apex of petrous bone. The cavernous sinus is reached. The dura is opened as in the retrolabyrinthine approach. If necessary, the trigeminal nerve may be mobilized by opening Meckel’s cave so that retraction of this nerve can be facilitated. The trigeminal nerve arises from the anterolateral surface of the pons. The sensory root is larger and the motor root is smaller. The motor root is medial to the sensory root. The nerve traverses the upper part of the cerebellopontine cistern and comes in close contact with the superior petrosal vein. The nerve enters Meckel’s cave below the superior petrosal sinus by pushing a layer of dura along with it. The dura blends with the epineurium at the level of the gasserian ganglion. From the ganglion the three branches arise. The motor root joins with the mandibular division.
Translabyrinthine Approach
After the mastoidectomy the labyrinthine procedure is performed. It involves removal of the semicircular canals and vestibule to expose the dura lining the internal auditory canal. After drilling and removing the SCCs, the vestibule is reached and removed. Care is needed to avoid injury to the facial nerve as it courses below the lateral SCC and the ampulla of the posterior SCC, and around the superolateral margin of the vestibule. Bone is further removed at the lateral end of the canal to expose the transverse and the vertical crest along with the four nerves at the lateral end. While removing the bone behind the posterior wall of the IAC, it is important to remember that the jugular bulb may bulge upward behind the posterior SCC or IAC. The vestibular aqueduct and endolymphatic sac are opened and removed as bone is removed between the meatus and the jugular bulb. The cochlear canaliculus is seen deep to the vestibular aqueduct as the bone is removed in between the jugular bulb and the IAM. The lower end of the cochlear canaliculus is situated just above the area where the glossopharyngeal nerve enters the medial side of the jugular foramen. Sometimes the subarcuate artery or even the AICA may be encountered in Trautmann’s triangle.
Total Petrosectomy Approach17
This approach is reserved for giant tumors or aneurysms in the clival-petroclival region. It is also suitable for patients who were previously operated and radiated. After making a wide C-shaped skin incision extending forward to the anterior aspect of the superior temporal line, the external auditory canal is transected and over-sewn. The temporomandibular joint capsule is dissected free from the glenoid fossa. A complete mastoidectomy and a labyrinthectomy are done. The facial nerve is exposed from the stylomastoid foramen to the IAC. A temporal craniotomy is done with an anterior extent up to the sphenoid wing. A zygomatic osteotomy including the condylar fossa is done to gain access to the vertical segment of the petrous carotid artery. Occasionally, for greater exposure, the mandibular condyle needs to be resected. Next, the GSPN and the MMA are cut. The cartilaginous eustachian tube is exposed, cut, and sewn. The petrous carotid artery is completely mobilized from the proximal cavernous sinus portion to the upper cervical segment, as mentioned previously. The fibrocartilaginous ring surrounding the cervical carotid at its entrance into the skull base is divided, and the ICA is mobilized anteriorly. The cochlea and the bone medial to the mastoid part of the facial nerve are removed. The jugular foramen and its associated structures are skeletonized, and the facial nerve is mobilized posteriorly. Next, the medial petrous apex and clivus can be resected.
Far Lateral Approach to the Foramen Magnum and the Craniovertebral Junction
This approach is used to access anterior or anterolateral lesions in the lower clivus, C1, and C2. A lazy S-shaped, straight, or an inverted U-shaped incision is used. The advantage of an inverted U-shaped incision is the ease of doing a fixation if necessary. Some surgeons cut through the muscle in layers; others prefer an anatomic dissection, detaching the muscles from their attachments in layers. The advantage of the anatomic dissection is the ease of identifying the vertebral artery, which is located below the deepest layer of the superior and inferior oblique muscles. These muscles are detached, and the vertebral artery covered by the venous plexus is identified. This step must be done under the microscope. The foramen transversarium of the atlas and often the axis are unroofed to expose the vertebral artery. The venous sheath is coagulated and slit open longitudinally along the course of the artery, and the artery is completely taken out of the sheath. Considerable venous bleeding is encountered during this step, but once the artery is removed from the sheath, the venous bleeding is easily controlled. The vertebral artery is then followed up to the intradural entry. The venous sheath is adherent to the capsule of the joint formed by the occipital condyle and the C1 lateral mass, and the vertebral artery must be carefully detached to mobilize it completely. During this procedure, it must be remembered that the PICA may have extradural origin. The mobilization and proximal control depends on the pathology and the available intradural length of the vertebral artery. The suboccipital bone, the posterior part of the mastoid, and the ipsilateral posterior arch of the atlas are exposed. A suboccipital craniotomy is performed to include the foramen magnum. The posterior border of the sigmoid sinus is exposed. Below the jugular bulb, the bone removal is taken up to the occipital condyle. The ipsilateral posterior arch of C1 is also removed up to the lateral mass. The removal of the atlas depends on the lower extent of exposure needed. Up to this point, the exposure is adequate for the lesions in the anterolateral aspect of the foramen magnum. For the lesions placed more anteriorly, further bone removal is necessary. The posterior one third of the occipital condyle and superiorly the jugular tubercle are drilled away. For tumors involving the occipital condyle, drilling of the entire condyle is necessary. In this case, stabilization is necessary.
After dural opening, the vertebral artery, dentate ligament, and cranial nerves, apart from the cerebellar tonsil and brainstem, come into view. The dentate ligament is a white fibrous sheet attached to the spinal cord medially and the dura laterally. Its medial border has a continuous linear attachment to the cord midway between the dorsal and ventral roots. Its lateral border is attached at intervals by fibrous triangular processes to the dura. The rostral-most process is attached to the dura at the foramen magnum level, and the caudal process is attached posterior and inferior to the initial intradural segment of the vertebral artery. The lateral border of the dentate ligament between the two rostral-most triangular processes is attached to the vertebral and posterior spinal arteries and to the C1 nerve roots, making separation of these structures difficult.
The rootlets of the hypoglossal nerve arise form the medulla along a line that is continuous with the line along which the ventral spinal roots arise. This line in the medulla is between the pyramid and the olive. These rootlets pass anterior to the vertebral artery and exit the intracranial compartment through the hypoglossal canal along with the venae comitans. The glossopharyngeal, vagus, and accessory nerves arise in a series of rootlets along a line posterior to the olive. They exit the intracranial compartment through the jugular foramen. At this point, where the glossopharyngeal nerve penetrates the dura, it is separated from the vagus nerve by a dural septum.1 The accessory nerve has two components. The cranial part is composed of rootlets arising from the medulla and the upper rootlets joining the vagus nerve. The spinal part is formed by the union of a series of rootlets, which arise from the lower medulla and upper spinal cord. The spinal component passes through the foramen magnum and is joined by the lower-most rootlets of the cranial component. The spinal component arises as a series of rootlets situated midway between the ventral and dorsal roots of spinal nerves. The rootlets contributing to the formation of the spinal accessory may arise as low as the C7 root level.
Approach to the Jugular Foramen
The jugular foramen area is one of the most complex areas to reach. The choice of surgical approach depends on the exact location of the lesion and the presence of vascular encasement.
Transjugular/Mastoid and Neck Approach
This approach is used for lesions in the jugular bulb without extension into the neck or posterior fossa. A curved postauricular incision is made. The skin incision is carried down to the neck along the anterior border of the sternocleidomastoid muscle, and the sternocleidomastoid muscle is detached from the mastoid and reflected posteriorly. The digastric muscle is detached from the digastric groove and is reflected anteriorly. A complete mastoidectomy is performed, and the mastoid tip is transected lateral to the digastric groove. This exposes the major neurovascular bundle. The spinal accessory nerve is identified crossing the internal jugular vein (IJV) lateral to it. The glossopharyngeal and vagus nerves are also identified and preserved. For paragangliomas, the IJV is doubly ligated in the neck. Next, the bone over the sigmoid sinus and the jugular bulb are completely drilled with a diamond bur. This exposes the entire jugular foramen region.
Mastoid and Neck Approach with Limited Facial Nerve Rerouting
This approach is needed for approaching more superiorly located lesions. Here, in addition to the approach described above, a total mastoidectomy is performed. The infralabyrinthine and retrofacial air cells are drilled to expose the facial nerve from the second genu to the stylomastoid foramen. The periosteum of the facial nerve canal is preserved, which in turn preserves the vascular supply of the facial nerve. The fibrous attachment at the stylomastoid foramen is sharply cut to mobilize the facial nerve anteriorly and laterally. This facilitates access to the jugular bulb. This approach is suitable for jugular foramen schwannomas and paragangliomas limited to the jugular bulb.
Infratemporal Approach18
This approach is suitable for paragangliomas intimately involving the carotid artery. A C-shaped incision is made about 2 inches behind the postauricular crease. It extends upward and forward along the superior temporal line and extends below along the anterior margin of the sternocleidomastoid to the level of the thyroid cartilage. The skin and the galeal flap are raised. The posterior wall of the external auditory canal is identified. Dissection of cervical flap is performed deep to the platysma muscle. The external auditory canal is transected at the level of the bony-cartilaginous junction. The cartilage is removed from the ear canal and the skin is fashioned as a cuff ready to be everted and sutured. A mastoidectomy is performed. The chorda tympani is sacrificed to widen the facial recess. The posterior wall of the external auditory canal is drilled and removed. The ossicles of the middle ear, tympanic membrane, and the remaining skin of the external auditory canal are removed. The facial nerve is exposed from the geniculate ganglion to the stylomastoid foramen level. Next, the neck is exposed to take proximal control. The tympanic ring is removed to expose the jugular bulb completely. The tympanic bone is then drilled along with the bone in the region of the temporomandibular joint to expose the petrous carotid artery. The facial nerve is transposed anteriorly and laterally. The rest of the bone covering the petrous carotid artery and jugular bulb is drilled to skeletonize all the neurovascular structures in the jugular foramen area.
Extended Subfrontal Approach to the Clivus and Foramen Magnum
A bicoronal skin flap is raised and the skin is reflected downward, exposing the orbital rim completely on both sides and beyond the frontonasal suture in the midline. The temporalis muscles are detached from the anterior temporal fossa and are retracted posteriorly. The periorbita is dissected from the roof and the upper medial wall of the orbit. A bifrontal craniotomy is performed. A bilateral orbital osteotomy is performed including the ethmoid. The orbito-fronto-ethmoid segment is loosened with an osteotome and removed after the attachments of the nasal mucosa and septum have been divided. The nasal cavity, ethmoid sinus, and orbits come into view. The ethmoid and nasal mucosa are vascular and should be cauterized. The posterior ethmoidal artery must be identified and cauterized. This is an important landmark, as the optic canal is just behind it. The optic canals may have to be decompressed depending on the pathology. The ethmoid cells are removed completely, which exposes the sphenoid sinus. The planum sphenoidale is resected. Then the sellar floor and lateral wall are drilled, completely exposing the cavernous sinus. Although it is difficult to drill the dorsum sellae, especially if it is prominent, it can be done with the help of a 30-degree endoscope. The medial cavernous sinus is protected by a thin periosteal layer, which may become torn or may have to be opened intentionally. Bleeding from the cavernous sinus is controlled with packing Surgicel or Gelfoam. The intracavernous ICA is followed along its anterior bend and medial surface of the horizontal and vertical segments to the petrocavernous junction. The medial part of the petrous apex as well as the body of the sphenoid bone may be removed this way. The clivus is drilled next, which exposes the clival dura. This may cause brisk bleeding from the basilar venous plexus. In young adults the dura is thick, but in the elderly it may be very thin and may be breached during drilling. The lateral boundaries of this approach are the cavernous sinuses, petrous apices, abducens nerves, and the hypoglossal canals. The lower boundary is the foramen magnum.
References
1. Katsuta T, Rhoton AL Jr, Matsushima T. The jugular foramen: microsurgical anatomy and operative approaches. Neurosurgery 1997;41:149–201, discussion 201–202 PubMed
2. Pait TG, Harris FS, Paullus WS, Rhoton AL Jr. Microsurgical anatomy and dissection of the temporal bone. Surg Neurol 1977;8:363–391 PubMed
3. Tedeschi H, Rhoton AL Jr. Lateral approaches to the petroclival region. Surg Neurol 1994;41:180–216 PubMed
4. Sekhar LN, Raso J, Schessel DA. The presigmoid petrosal approach. In: Sekhar LN, Oliveira ED, eds. Cranial Microsurgery: Approaches and Techniques. New York: Thieme, 1999:432–463
5. Bricolo A, Turazzi S. Surgery for gliomas and other mass lesions of the brainstem. In: Symon L, Calliauw L, Cohadon, et al, eds. Advances and Technical Standards in Neurosurgery, Vol 22. Vienna: Springer-Verlag, 1995:261–341
6. Gortvai P, Teruchkin S. The position and extent of the human dentate nucleus. Acta Neurochir (Wien) 1974;21(Suppl 21):101–110 PubMed
7. Lister JR, Rhoton AL Jr, Matsushima T, Peace DA. Microsurgical anatomy of the posterior inferior cerebellar artery. Neurosurgery 1982;10:170–199 PubMed
8. Macchi V, Porzionato A, De Caro R, Clemente A, Parenti A. Comment on “the relationship of the posterior inferior cerebellar artery to the cranial nerves VII-XII.” Clin Anat 2008;21:218–220, author reply 221 PubMed
9. Fine AD, Cardoso A, Rhoton AL Jr. Microsurgical anatomy of the extracranial-extradural origin of the posterior inferior cerebellar artery. J Neurosurg 1999;91:645–652 PubMed
10. Mussi ACM, Rhoton AL Jr. Telovelar approach to the fourth ventricle: microsurgical anatomy. J Neurosurg 2000;92:812–823 PubMed
11. Avci E, Chanda A, Fossett DT. Preauricular subtemporal infratemporal approach. In: Fossett DT, Caputy AJ, eds. Operative Neurosurgical Anatomy. New York: Thieme, 2002:52–60
12. Sen CN, Sekhar LN. The subtemporal and preauricular infratemporal approach to intradural structures ventral to the brain stem. J Neurosurg 1990;73:345–354 PubMed
13. Rhoton AL Jr. Microsurgical anatomy of the brainstem surface facing an acoustic neuroma. Surg Neurol 1986;25:326–339 PubMed
14. Rhoton AL Jr, Kobayashi S, Hollinshead WH. Nervus intermedius. J Neurosurg 1968;29:609–618 PubMed
15. Chanda A, Nanda A. Partial labyrinthectomy petrous apicectomy approach to the petroclival region: an anatomic and technical study. Neurosurgery 2002;51:147–159, discussion 159–160 PubMed
16. Sekhar LN, Schessel DA, Bucur SD, Raso JL, Wright DC. Partial labyrinthectomy petrous apicectomy approach to neoplastic and vascular lesions of the petroclival area. Neurosurgery 1999;44:537–550, discussion 550–552 PubMed
17. Cass SP, Sekhar LN, Pomeranz S, Hirsch BE, Snyderman CH. Excision of petroclival tumors by a total petrosectomy approach. Am J Otol 1994; 15:474–484 PubMed
18. Fisch U. Infratemporal fossa approach for glomus tumors of the temporal bone. Ann Otol Rhinol Laryngol 1982;91(5 Pt 1):474–479 PubMed
Related posts:
 Shunt Surgery for Posterior Fossa Lesions
Shunt Surgery for Posterior Fossa Lesions
 The Retromastoid Approach: Techniques and Results
The Retromastoid Approach: Techniques and Results
 Posterior Fossa Arteriovenous Malformations and Cavernous Angiomas
Posterior Fossa Arteriovenous Malformations and Cavernous Angiomas
 Neuroimaging of the Posterior Fossa
Neuroimaging of the Posterior Fossa
 Microvascular Decompressions for Hemifacial Spasm and Trigeminal Neuralgia
Microvascular Decompressions for Hemifacial Spasm and Trigeminal Neuralgia
 Aneurysms of the Basilar Trunk: Far Lateral and Petrosal Approaches
Aneurysms of the Basilar Trunk: Far Lateral and Petrosal Approaches
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


