Chapter 23 Muscle
Skeletal Muscle
Shape and Fibre Architecture
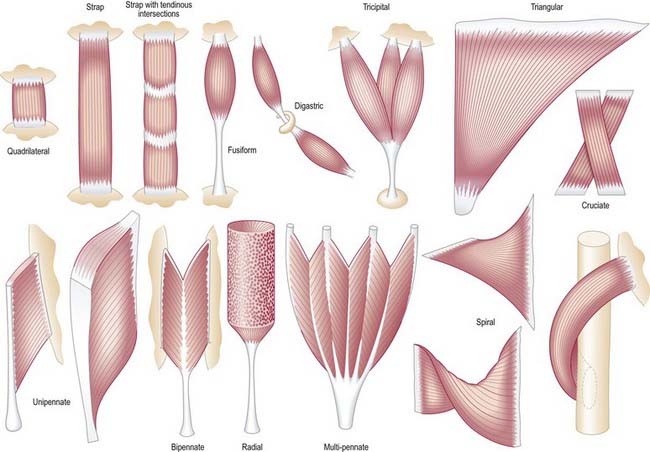
It is possible to classify muscles based on their general shape and the predominant orientation of their fibres relative to the direction of pull (Fig. 23.1). Muscles with fibres that are largely parallel to the line of pull vary in form from flat, short and quadrilateral (e.g. thyrohyoid) to long and strap-like (e.g. sternohyoid, sartorius). In such muscles, individual fibres may run for the entire length of the muscle or over shorter segments when there are transverse, tendinous intersections at intervals (e.g. rectus abdominis). In a fusiform muscle, the fibres may be close to parallel in the belly but converge to a tendon at one or both ends. Where fibres are oblique to the line of pull, muscles may be triangular (e.g. temporalis, adductor longus) or pennate (feather-like) in construction. The latter vary in complexity (see Fig. 23.1) from unipennate (e.g. flexor pollicis longus) and bipennate (e.g. rectus femoris, dorsal interossei) to multipennate (e.g. deltoid). In some muscles, the fibres pass obliquely between deep and superficial aponeuroses, in a type of unipennate form (e.g. soleus). In other sites, muscle fibres start from the walls of osteofascial compartments and converge obliquely on a central tendon in circumpennate fashion (e.g. tibialis anterior). Some muscles have a spiral or twisted arrangement (e.g. sternocostal fibres of pectoralis major and latissimus dorsi, which undergo a 180° twist between their median and lateral attachments). Others spiral around a bone (e.g. supinator, which winds obliquely around the proximal radial shaft) or contain two or more planes of fibres arranged in different directions, a type of spiral sometimes referred to as cruciate; sternocleidomastoid, masseter and adductor magnus are all partially spiral and cruciate. Many muscles display more than one of these major types of arrangement and show regional variations that correspond to contrasting and, in some cases, independent actions.
Muscle Nomenclature
The names given to individual muscles are usually descriptive, based on their shape, size, number of heads or bellies, position, depth, attachments or actions. The meanings of some of these terms are given in Table 23.1.
Table 23.1 Terms used in naming muscles
| Shape |
| Size |
| Number of Heads or Bellies |
| Depth |
| Position |
| Attachment |
| Action |
Microstructure
The cellular units of skeletal muscle are the muscle fibres (Fig. 23.2). These long, cylindrical structures tend to be consistent in size within a given muscle, but in different muscles they may range from 10 to 100 µm in diameter and from millimetres to many centimetres in length. Some typical skeletal muscle fibres are seen in longitudinal section in Figure 23.3. Their staining characteristics are dominated by the contractile apparatus, which constitutes much of the cytoplasm or sarcoplasm. The contractile proteins are organized into cylindrical myofibrils that are too tightly packed to be visible by routine light microscopy. Of greater significance are transverse striations, which are the result of alignment across the fibre of repeating elements, the sarcomeres, within neighbouring myofibrils. These cross-striations are usually evident in sections stained conventionally, but they may be demonstrated more effectively using special stains (Fig. 23.4).
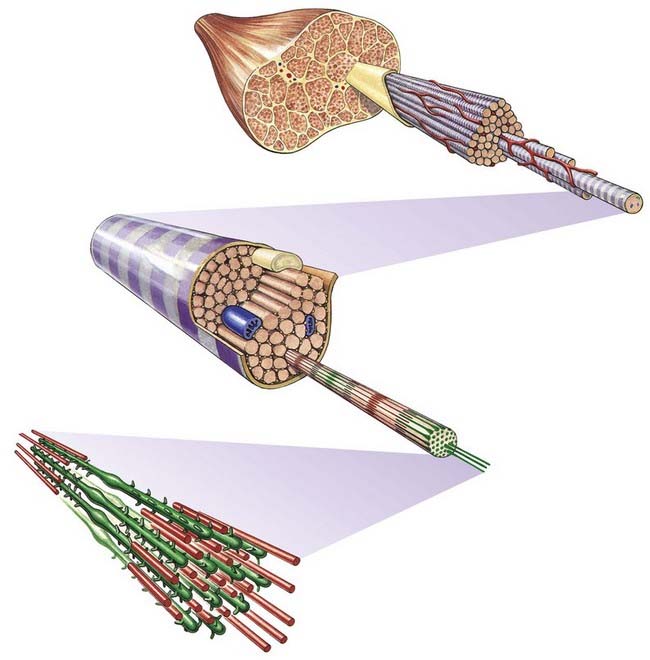
Fig. 23.2 Levels of organization within a skeletal muscle, from whole muscle to fasciculi, single fibres, myofibrils and myofilaments.
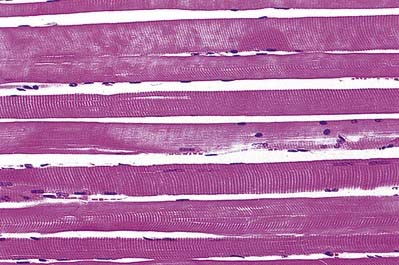

Under polarized light, the striations are even more striking and are seen as a pattern of alternating dark and light bands. The darker bands are birefringent, rotating the plane of polarized light strongly, and are known as anisotropic or A-bands; the lighter bands rotate the plane of polarized light to a negligible degree and are known as isotropic or I-bands. The structures responsible for this appearance are described more readily at the ultrastructural level.
The multiple nuclei are oval and are located at the periphery of the fibres, under the plasma membrane or sarcolemma. They are especially numerous in the region of the neuromuscular junction. The nuclei are moderately euchromatic and usually have one or more nucleoli. They occupy a thin, transparent rim of sarcoplasm between the myofibrils and the sarcolemma and are seen most clearly in transverse sections (Fig. 23.5). Other nuclei belonging to vascular endothelial cells, Schwann cells, fibroblasts, and so forth may be present in the spaces between the fibres, where blood vessels and nerve fibres travel through layers of fine connective tissue, the endomysium. Nuclei of satellite cells lie between the sarcolemma and the surrounding basal lamina.
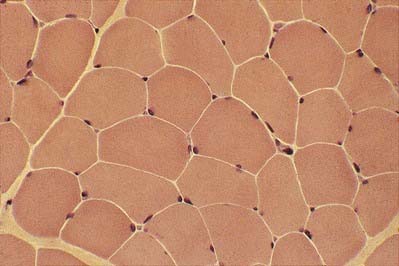
In transverse section, the profiles of the fibres are usually polygonal (see Fig. 23.5). Some muscles, such as the extrinsic muscles of the larynx, tend to be less tightly packed. In such situations, as well as in conditions of generalized wasting or muscle damage, the fibres may adopt a more rounded profile; however, in some normal muscles, such as those that close the jaw, the fibres are closely packed but have rounded profiles. The sarcoplasm often has a stippled appearance because the transversely sectioned myofibrils are resolved as dots.
Skeletal muscle fibres are large (with a few exceptions, such as the laryngeal muscles), and unless electron micrographs are taken at very low magnification, they seldom show more than part of the interior of a fibre (Fig. 23.6A). Myofibrils are the dominant ultrastructural feature of such micrographs. They are cylindrical structures approximately 1 µm in diameter, which appear as ribbons in longitudinal section. Thin, very densely stained transverse lines, which correspond to discs in the parent cylindrical structure, appear at regular intervals along these ribbons. They are called Z-lines or, more properly, Z-discs (Zwischenscheiben, or ‘between discs’). They divide the myofibril into a linear series of identical contractile units called sarcomeres, each of which is approximately 2.2 µm long in resting muscle.
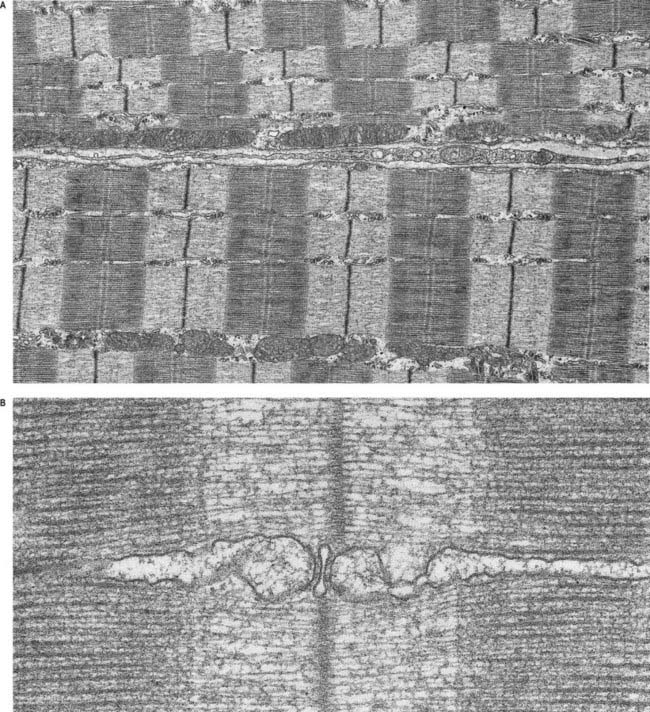
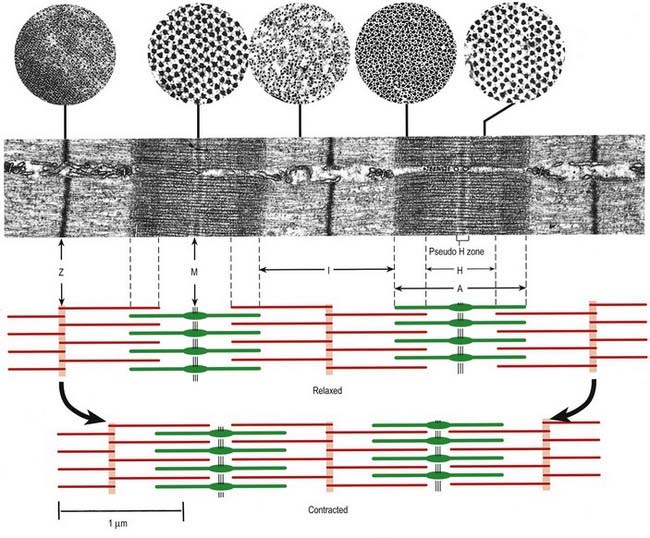
At higher power, sarcomeres are seen to consist of two types of filament—thick and thin—organized into regular arrays (Fig. 23.6B). The thick filaments, which are approximately 15 nm in diameter, are composed mainly of myosin. The thin filaments, which are 8 nm in diameter, are composed mainly of actin. The arrays of thick and thin filaments form a partially overlapping structure in which the electron density varies according to the amount of protein present. The A-band consists of the thick filaments, together with lengths of thin filaments that interdigitate with, and thus overlap, the thick filaments at either end (see Fig. 23.6B; Fig. 23.7). The central, paler region of the A-band, into which the thin filaments have not penetrated, is called the H zone (helle, or ‘light’). At their centres, the thick filaments are linked together transversely by material that constitutes the M line (Mittelscheibe, or ‘middle (of) disc’), which is visible in most muscles.






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


