19 Vestibular nerve
Introduction
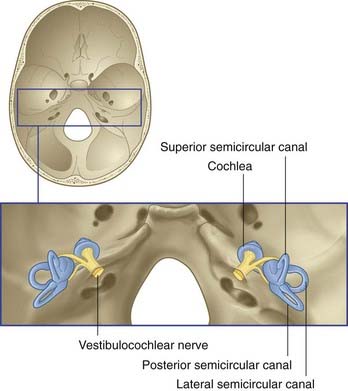
The vestibulocochlear nerve is primarily composed of the centrally directed axons of bipolar neurons housed in the petrous temporal bone (Figure 19.1). The peripheral processes are applied to neuroepithelial cells in the vestibular labyrinth and cochlea. The nerve enters the brainstem at the junctional region of pons and medulla oblongata. The functional anatomy of the vestibular division of the nerve is described in this chapter.
Vestibular System
The vestibular labyrinth comprises the utricle, the saccule, and the cristae within three semicircular ducts (Figure 19.2). The utricle and saccule contain a 3 × 2 mm2 macula. Each semicircular duct contains an ampulla at one end, and the ampulla houses a crista. (It should be pointed out that clinicians commonly speak of ‘canals’ where ‘ducts’ would be strictly more appropriate.)
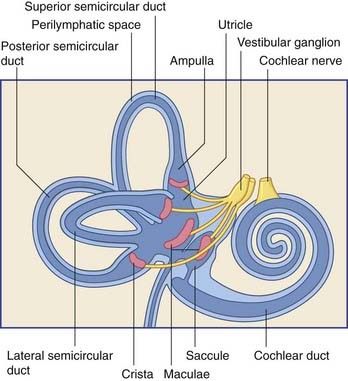
The bipolar cells of the vestibular (Scarpa’s) ganglion occupy the internal acoustic meatus. Their peripheral processes are applied to the five sensory end organs. Their central processes, which constitute the vestibular nerve, cross the subarachnoid space and synapse in the vestibular nuclei previously seen in Figures 17.14 and 17.15.
Static labyrinth: anatomy and actions
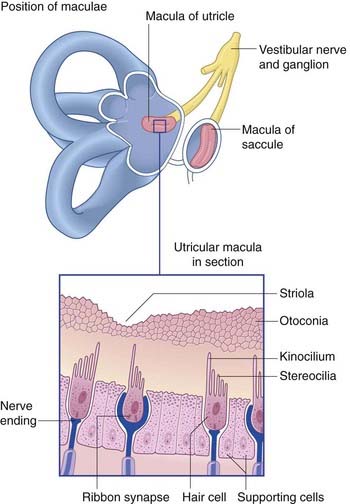
The position and structure of the maculae are shown in Figure 19.3. The utricular macula is relatively horizontal, the saccular macula is relatively vertical. The cuboidal cells lining the membranous labyrinth become columnar supporting cells in the maculae. Among the supporting cells are so-called hair cells, to which vestibular nerve endings are applied. Some hair cells are almost completely enclosed by large nerve endings, whereas others (phylogenetically older) receive only small contacts. At the cell bases are ribbon synapses, the synaptic vesicles being lined up along synaptic bars. Projecting from the free surface of each hair cell are about 100 stereocilia and, close to the cell margin, a single, long kinocilium. The hair cells discharge continuously, the resting rate being about 100 Hz.
The lateral vestibulospinal tract, seen earlier in sections of medulla oblongata in Chapter 17, arises from large neurons in the lateral vestibular nucleus (of Deiters). The fibers descend in the anterior funiculus on the same side of the spinal cord and synapse upon extensor (antigravity) motor neurons. Both α and γ motor neurons are excited, and a significant part of the increased muscle tone is exerted by way of the gamma loop (Ch. 16). During standing, the tract is tonically active on both sides of the spinal cord. During walking, activity is selective for the quadriceps motor neurons of the leading leg; this commences following heel strike and continues during the stance phase (when the other leg is off the ground). Deiters’ nucleus is somatotopically organized, and the functionally appropriate neurons are selected by the flocculonodular lobe of the cerebellum. The flocculonodular lobe (Ch. 25) has two-way connections with all four vestibular nuclei.






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


