connections; thus, each brainstem nucleus as well as the cortex has an input from both ears. As shown in Figure 47.1, this functional organization has been demonstrated in humans with recording of EPs during surgical procedures for the treatment of intractable epilepsy (1, 2 and 3).
Table 47.1 Classification of Auditory Evoked Potentials According to Latency | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
abruptly by a larger negative wave, the AP or N1 potential. The CM represents (Table 47.1) the alternating current (AC) component of the hair cell receptor potential (29, 30 and 31), or the fluid pressure (condensation and rarefaction) produced in the cochlear fluid by sound-induced stapes footplate vibrations (32); SP represents the direct current (DC) potentials mostly from inner hair cells of the organ of Corti of the basal portion of the cochlea with some contribution from the outer inner hair cells (27,30,31). The AP represents the action potential of the auditory nerve and therefore is the same potential as Wave I of the BAEPs. AP (N1) is usually larger than the BAEPs Wave I, because of the methodology of recording the potentials closer to the nerve at the tympanic membrane. Thus, ECochG is employed when less than optimal recording conditions were used to obtain Wave I of the BAEPs (27). ECochG recording can assess the functional integrity of the cochlea and the auditory nerve and in diseases affecting the hearing system determine what is the contribution of each of these structures to the patient deficit (27,28,33, 34, 35 and 36).
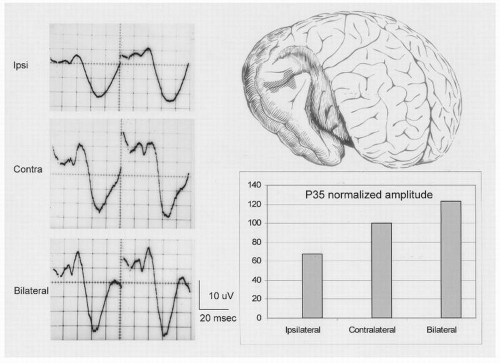 Figure 47.1 Auditory evoked responses recorded directly from the human cortex (area in hatching in the brain drawing on the upper right of the figure) exposed during temporal lobectomy for the treatment of medically intractable epilepsy. Responses from monocular stimulation of the ipsilateral ear are smaller than responses from stimulation of the contralateral ear. Bilateral ear stimulation results in the highest amplitude responses. Each potential is the average of 64 responses; positive deflections are downward. The evoked responses are shown on the left half of the figure; on the right half is a bar graph of the amplitude of the P35 to the three different stimuli. |
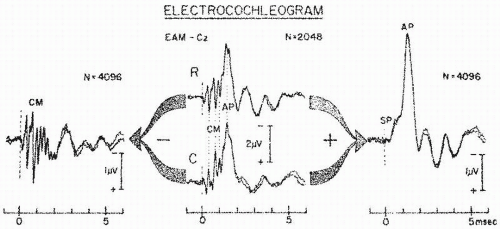 Figure 47.2 Electrocochleogram elicited by exclusively rarefaction (R) or condensation (C) clicks (middle section) are composites of CM, SP, and AP potentials. Dotted line emphasizes reversal of CM polarity and lack of AP polarity reversal when stimulus phase is inverted. SP polarity behaves similarly to that of AP. Algebraically subtracting C from R ECochGs results in summation of CM and cancellation of most of the SP and AP (left), whereas adding C to R ECochGs results in summation of SP and AP and cancellation of most CM (right). (From Chatrian G, Wirch AL, Lettich K, et al. Click-evoked human electrocochleogram. Noninvasive recording method, origin and physiologic significance. Am J EEG Technol. 1982;22:151-174.) |
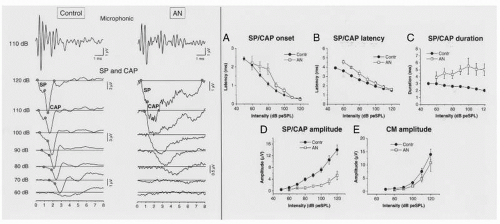 Figure 47.3 The left half of the figure represents the receptor potentials (CM and SP) and compound action potentials (CAP) obtained from both a control and one auditory neuropathy (AN) patient. The upper panel reports the cochlear microphonics recorded at 110 dB pe SPL at the same gain in control and AN subject. The lower panel reports the CAP with the superimposed SP at decreasing stimulation intensities from 120 to 60 dB pe SPL. Note the decrease in amplitude and the broadening in duration of the neural response obtained from AN subject compared to control. At threshold, the ECochG potentials from both AN and control have similar form. The right half of the figure represents the means and standard errors of SP/CAP onset (A), peak latency (B), duration (C), peak amplitude (D), and CM amplitude (E) for controls (filled circles) and AN subjects (open squares). There are significant differences between the two groups for SP/CAP peal latency, duration and peak amplitude. (Modified from Santarelli R, Starr A, Michalewski HJ, et al. Neural and receptor cochlear potentials obtained by transtympanic electrocochleography in auditory neuropathy. Clin Neurophysiol. 2008;119:1028-1041.) |
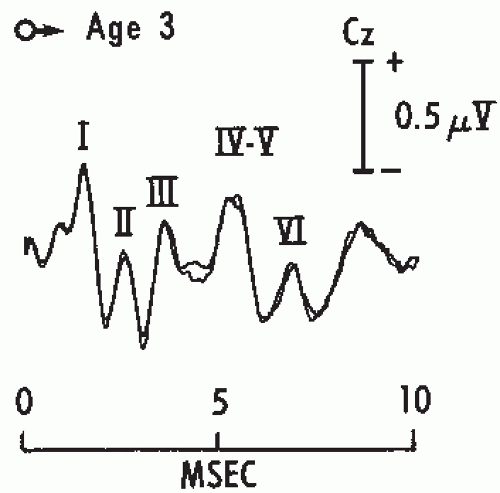 Figure 47.4 Brainstem auditory evoked potentials (BAEPs) in a normal child age 3. Two tracings were superimposed to show reproducibility of the responses. Recording was between Cz and the ipsilateral ear. Click intensity was 110 dB SPL. Note that Waves IV and V are fused together in a single complex. |
interaction of two dipoles, one originating in the ventral cochlear nucleus and the other in the ipsi- and contralateral trapezoid body and superior olivary complex. These data suggest that the origin of Wave III is within the brainstem (57).
Table 47.2 Origin of BAEP Waves | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
recordings to ipsi- and contralateral derivations are recommended to improve wave detection. Wave I (Fig. 47.5) is selectively attenuated in contralateral derivation, whereas Waves IV and V are separated better in contralateral than in ipsilateral recordings (73,74). The best way to obtain a large Wave I is to utilize a ball electrode placed in the external auditory canal (extratympanic electrode referred to the vertex [Fig. 47.6]); in this fashion it is feasible to simultaneously record ECochG and BAEPs.
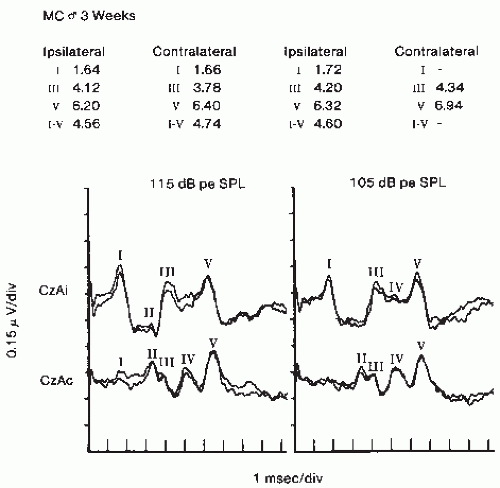 Figure 47.5 Brainstem auditory evoked potentials (BAEPs) in a 3-week-old boy. Note the greater amplitude of Wave I from ipsilateral recording but better definition of Wave IV in contralateral recording. CZ, vertex; Ai, ipsilateral earlobe; Ac, contralateral earlobe. Responses on the left half of the figure were to clicks of 115 dB SPL; responses on the right half of the figure were to clicks of 105 dB SPL. |
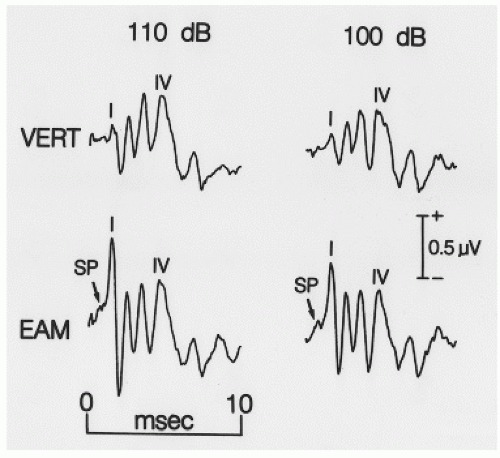 Figure 47.6 Simultaneous recordings of evoked potentials to monoaural clicks at 110 and 100 dB pe SPL. VERT indicates recordings from ipsilateral mastoid electrode referred to Cz; EAM indicates recordings from ipsilateral extratympanic electrode referred to Cz. Note that the recording from EAM shows a larger Wave I and also the summating potential (SP) of ECochG. |
cells, inner hair cells, and outer hair cells in 21 humans with presbycusis and noted that the extent of degeneration was associated with the severity of hearing loss. The possibility that some changes in the central auditory pathways contribute to the age-related delays of BAEPs cannot be entirely excluded. There have been reports of reductions in the number of neurons in the cochlear nucleus in aged humans (96,97). Gender also influences the various BAEP waves; there are consistently shorter latencies in females (73,80,81,92). Male-female differences are attributed to different body and brain size.
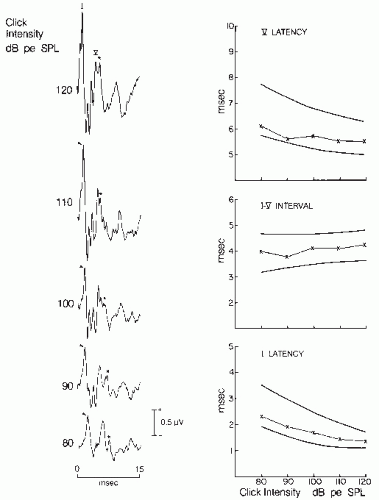 Figure 47.7 Brainstem auditory evoked potentials (BAEPs) in a normal 32-year-old woman. Results to monaural stimulation of the left ear. Recordings were obtained with a plastic-leaf silver electrode placed in the external auditory meatus and referred to the vertex. BAEPs to clicks of decreasing intensity are illustrated (left). Waves I and V are labeled and tagged with an arrow and an asterisk. Note the SP response preceding Wave I. The latency-intensity functions of Waves I and V, and I to V interval are shown (right). The thick lines represent the boundary of normal. Note that the value of the subject marked by an X is well within the normal range. pe, peak equivalent; SPL, sound pressure level. |
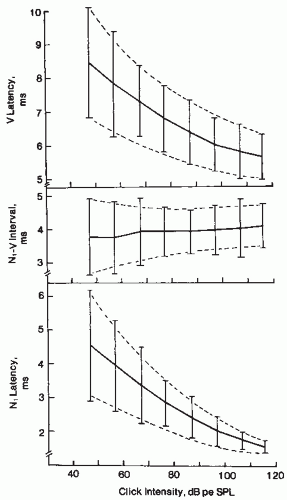 Figure 47.8 Latency-intensity curves from 23 normal-hearing ears showing means (solid lines) and ±2 standard deviation limits (crossbar). Dashed lines show estimated normal ranges. (From Coats A. Human auditory nerve action potentials and brain-stem evoked responses latency/intensity functions in detection of cochlear and retrocochlear pathology. Arch Otolaryngol. 1978;105:709-717.) |
vertex in such cases usually reveals a normal Wave I. Second, absence of Wave III is significant only when Wave V is also missing or delayed. Third, BAEPs cannot be interpreted without considering the patient’s hearing status; conductive hearing loss and cochlear pathology may profoundly affect BAEP wave latency and amplitude (12,23,63,86).
Latency-intensity functions indicating conductive hearing loss (Fig. 47.9).
The functions are characterized by prolonged Waves I and V with latency-intensity curves parallel to the normal curve. The I to V and I to III intervals are normal.
Latency-intensity functions indicating cochlear hearing loss (Fig. 47.9).
This type of abnormality accompanies high-frequency hearing loss of cochlear origin. It is characterized by a recruiting curve for Wave I, that is, normal or mildly prolonged Wave I latencies with loud clicks and greater delays with decreased intensity, resulting in a steep curve. Wave V is not drastically affected, and its curve is less steep, resulting in a shortened I to V interval.
Latency-intensity functions indicating retrocochlear deficit Type I (Fig. 47.10).
Wave I is prolonged with a steep latency-intensity function; since Wave V is prolonged, the I to V interval is prolonged. This type of abnormality has been reported in lesions affecting the eighth nerve.
Latency-intensity functions indicating retrocochlear deficit Type II (Fig. 47.10).
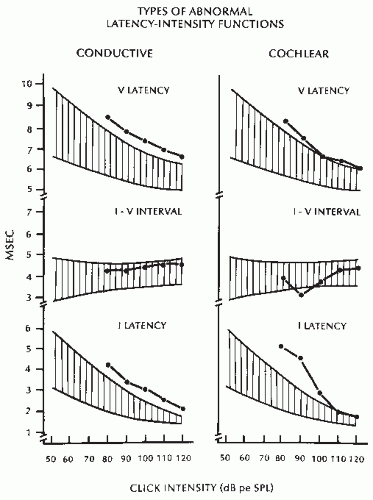 Figure 47.9 Latency-intensity functions in conductive and cochlear hearing loss. The hatched area represents the boundary of normality. The line connecting the dots represents the latency-intensity function for the two types of abnormalities. (See text for details.) |
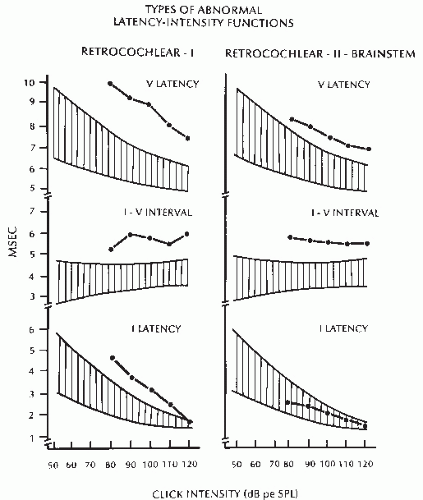 Figure 47.10 Two types of latency-intensity functions in retrocochlear dysfunction. (See text for details.) |
indicates a deficit at or after the superior olivary complex, in either the high pons or the low midbrain (17,98, 99 and 100). This detailed interpretation is based on two assumptions—that each BAEP wave is generated in a known neuronal location and that detailed identification of a lesion within brainstem structures is important in diagnosis and management of neurologic disorders—neither of which is correct. There is no consensus on the origin of BAEPs. Furthermore, the clinician is interested only in whether the lesion is intra-axial (within the brainstem) or extra-axial (in the VIII nerve). BAEPs cannot specify the nature of the lesion but only the type and approximate location of dysfunction.
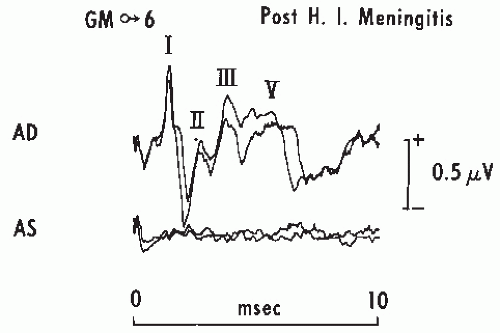 Figure 47.11 Brainstem auditory evoked potentials (BAEPs) in a boy, aged 6, 10 days after recovery from Haemophilus influenzae meningitis. Note the total absence of any response to left ear stimulation (AS), suggesting profound damage to the left cochlea or VIII nerve. Stimulation consisted of rarefaction clicks at 75 dB HL. Stimulation of the right ear AD shows a prolonged I to V interval and a dispersed Wave V amplitude suggesting right brainstem damage. |
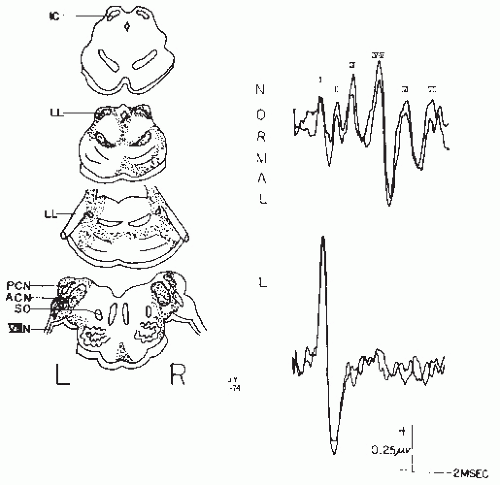 Figure 47.12 The distribution of neuropathology and BAEPs in a patient with anoxic brain damage. The BAEPs were recorded when neurologic examination and EEG were compatible with brain death. Note that the response to a 65 dB HL click consisted of only prolonged latency Wave I, compared to the normal record. (From Starr A, Hamilton AE. Correlation between confirmed sites of neurologic lesions and abnormalities of far-field auditory brainstem responses. Electroencephalogr Clin Neurophysiol. 1976;41:595-608.) |
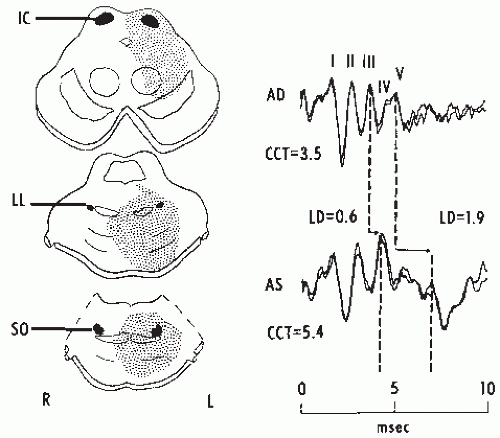 Figure 47.13 Brainstem auditory evoked potentials (BAEPs) in a male, aged 31, with a pontine glioma. The diagrams on the left show the patient’s lesion as seen by computed tomography the same day as the BAEP recordings. Note the progressively increased latencies of Waves III and V with stimulation of the left ear (AS), resulting in a prolonged I to V interval (CCT) of 5.4 msec. Wave V is also markedly depressed. The diagnosis was confirmed postmortem. IC, inferior colliculus; LL, lateral lemniscus; SO, superio-olive; AD, right ear; LD, latency difference. |
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


