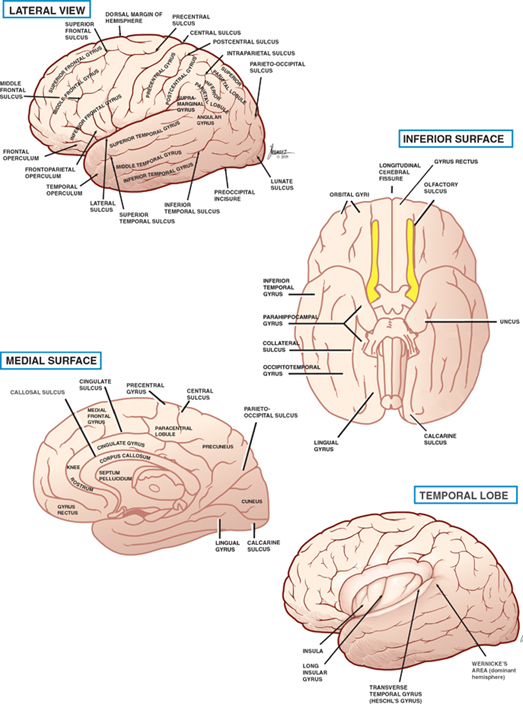
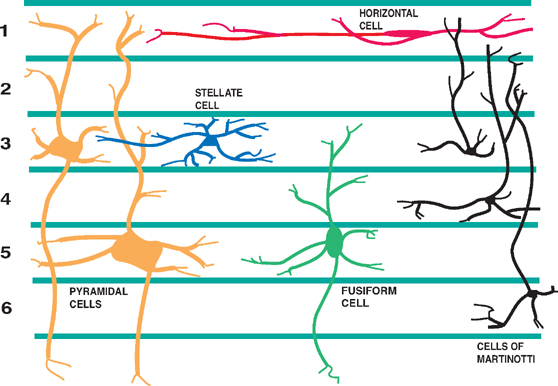
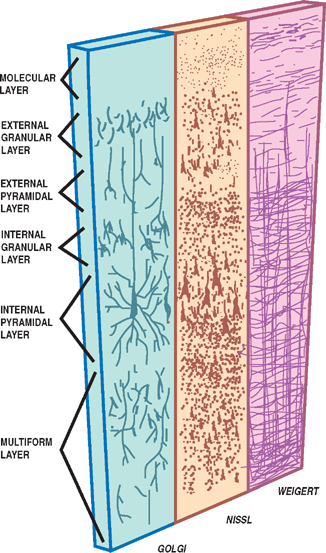
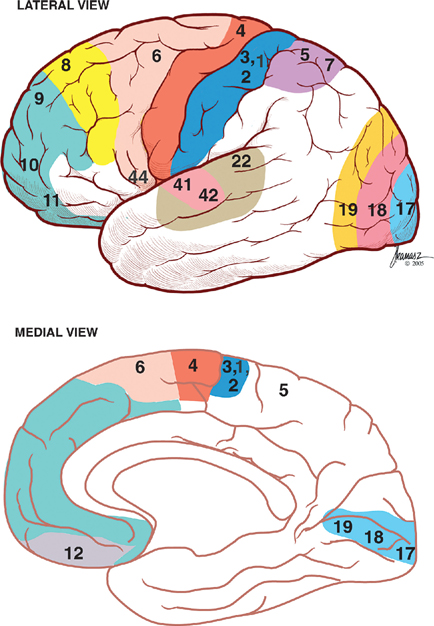
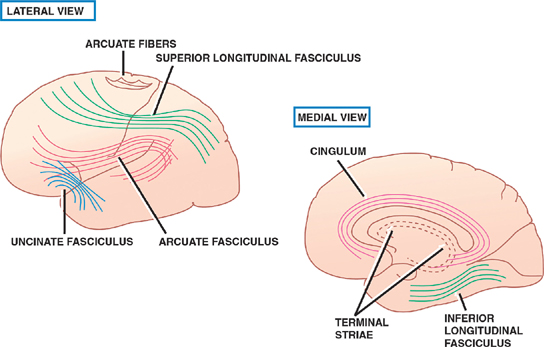
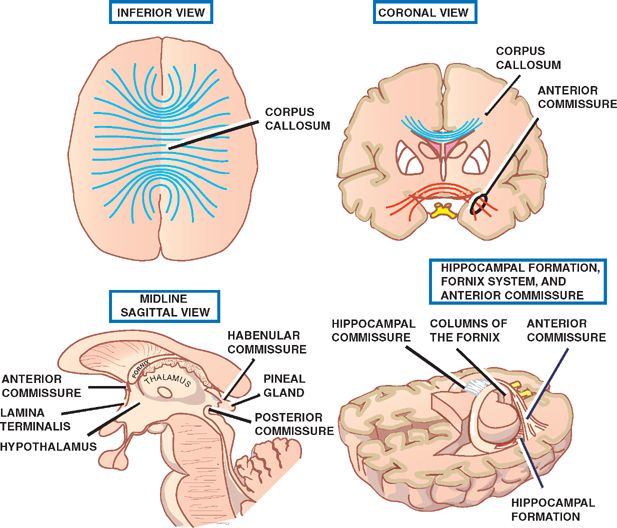
13 Cerebral Cortex See Fig. 13.1. The frontal, temporal, parietal, and occipital lobes are delimited by the lateral, parieto-occipital, central (rolan-dic), and calcarine sulci. The lateral sulcus, or sylvian fissure as it is also known, is the most constant fissure in the brain. It begins as a deep furrow on the inferior surface of the hemisphere and extends laterally between the frontoparietal and temporal lobes. The central sulcus vertically separates the frontoparietal cortices and, therefore, the motor (ventral) from the sensory (dorsal) cortex. It slopes downward from the cerebral convexity and forward 70 degrees to the horizontal plane and runs ~2 cm deep. The central sulcus is mainly a continuous sulcus. It rarely dips into the sylvian fissure. The walls of the calcarine sulcus, which lies on the medial surface of the occipital lobe, are formed by the visual cortex. It begins at the dorsal end of the corpus callosum and follows an arched course from the parieto-occipital sulcus to the occipital pole. The parieto-occipital sulcus extends vertically from the calcarine sulcus to the rostral border of the hemisphere to separate the parietal and occipital lobes. It begins ~4 cm from the occipital pole. The interhemispheric fissure divides the cerebrum into two hemispheres (right and left). The caudal border of the interhemispheric fissure is formed by the corpus callosum, a white matter tract that contains fibers that cross from one hemisphere to the other. The falx cerebri is a dural fold that extends into the interhemispheric fissure and serves to anatomically separate the right from the left cerebral hemispheres. The transverse cerebral fissure marks the separation between the cerebral hemispheres above and the cerebellum, midbrain, and diencephalon below. The dorsal portion of the transverse cerebral fissure intervenes between the occipital lobe above and the cerebellum below. It contains the tentorium cerebelli, a dural fold. The ventral aspect of the transverse cerebral fissure separates the corpus callosum above from the diencephalon below. It contains the tela choroidea, a vascular plexus derived from pia mater. Fig. 13.1 Major sulci and fissures. See Fig. 13.2. The precentral sulcus is discontinued and runs parallel to the central sulcus, just ventral to the precentral gyrus, which comprises the primary motor cortex. Ventral to the precentral sulcus are the superior, middle, and inferior frontal gyri, which are separated by the superior and inferior frontal sulci. In the dominant hemisphere, the posterior part of the inferior frontal gyrus contains Broca’s area, which controls expressive speech function. The postcentral gyrus runs parallel and dorsal to the central sulcus. It comprises the primary sensory cortex. The intraparietal sulcus, which extends from the postcentral sulcus dorsally, separates the dorsal parietal cortex into the superior and inferior parietal lobules. The inferior parietal lobule contains the supramarginal and angular gyri, which make up part of the receptive language area (Wernicke’s area) in the dominant hemisphere. The lateral surface of the temporal lobe comprises the superior, middle, and inferior temporal gyri, which are separated by the superior and inferior temporal sulci. The transverse temporal gyri (Heschl’s convolutions) lie adjacent to that portion of the superior temporal gyrus that forms the floor of the lateral sulcus. Heschl’s gyri are buried in the temporal operculum (within the insular cortex). They are not visualized on inspection of the lateral cerebral hemisphere. The cerebral hemispheres contain the primary auditory cortex on each side. The dorsal aspect of the superior temporal gyrus contains part of the receptive language region (Wernicke’s area) in the dominant hemisphere. Aside from the calcarine sulcus already described, the occipital cortex contains minor and extremely variable sulci that may rarely result in the definition of superior, middle, and inferior occipital gyri. The cingulate gyrus lies rostral to the corpus callosum along its entire course. The gyrus is separated from the corpus callosum by the sulcus of the corpus callosum (callosal sulcus) and the indusium griseum, a thin layer of cortical gray matter that lies on the superior surface of the corpus callosum. The paracentral lobule surrounds the indentation made by the central sulcus. It is formed by medial extensions of the precentral and postcentral gyri of the lateral hemisphere. The paracentral lobule contains parts of the primary motor and primary sensory cortices. In addition, the supplementary motor area lies in the anterior aspect of the paracentral lobule. The precuneus is located just dorsal to the paracentral lobule. It is formed by medial extensions of the superior parietal lobule of the lateral hemisphere. The cuneus is located in the occipital lobe just dorsal to the precuneus. The precuneus is separated from the cuneus by the parieto-occipital sulcus. A convolution on the inferior undersurface of the hemisphere extends from the occipital pole almost to the temporal pole. Ventrally, it contains the parahippocampal gyrus; dorsally, it contains the medial occipitotemporal gyrus. Deep to the parahippocampal gyrus is the hippocampus. The ventromedial extent of the parahippocampal gyrus contains the uncus. Herniation of the uncus as the result of increased intracranial pressure results in early oculomotor nerve palsy that may progress into a coma and cardiorespiratory compromise. Laterally, these gyri are bounded by the collateral sulcus. Just lateral to the collateral sulcus is the lateral occipitotemporal gyrus and the inferior part of the inferior temporal gyrus. The lingual gyrus extends forward into the temporal lobe, where it is continuous with the parahippocampal gyrus. The inferior frontal lobe is marked by the olfactory sulcus, which contains the olfactory bulb and tract. The olfactory sulcus is bordered medially by the gyrus rectus and laterally by the orbital gyri. Fig. 13.2 Gyri and sulci. See Fig. 13.3. A variety of cells make up the cerebral cortex. These include both neurons and glia. Glial cells, which include oligodendrocytes, astrocytes, and microglial cells, serve to support the neurons. Examples of these roles include providing metabolic support, scavenging neurotransmit-ters, and providing myelination. Neurons are the targets or cells of origin of all of the information traveling into and out of the cerebral cortex, respectively. The pyramidal cell neurons, which are the most numerous cells of the cerebral cortex, constitute about two thirds of the cortical neurons. The pyramidal cells communicate with other neurons in one of three ways: (1) projection neurons send axons out of the cerebral hemisphere to such areas of the brain as the corpus striatum, the brainstem, the spinal cord, and the thalamus; (2) association neurons connect to other neurons in the same hemisphere; and (3) commissural neurons send axons to the contralateral hemisphere. The most obvious example is the corpus callosum. Stellate cells are interneurons present in the cerebral cortex. They are smaller in size (compared with pyramidal cells), polygonal in shape, and possess multiple branching dendrites with a relatively short axon. They are the only excitatory interneurons in the cerebral cortex that use the neurotransmitter glutamate. All other cortical in-terneurons are gamma-aminobutyric acid (GABA)—containing inhibitory neurons. Fusiform cells are also interneurons and are concentrated mainly in the deepest layers of the cortex (i.e., farthest from the cortical surface). They send dendrites toward the surface of the cortex to branch and infuence neurons in the superficial cortical layers. The horizontal cells are confned to the most superf-cial layer of the cortex. They are small fusiform cells that are horizontally oriented. These axons run parallel to the surface of the cortex, making contact with the ascending dendrites of pyramidal cells. The cells of Martinotti are present at many levels of the cortex. They send axons that end in the most superficial layers of the cortex. Fig. 13.3 Cortical neurons. See Fig. 13.4. The thickness of the cerebral cortex varies from 1.5 to 4.5 mm. The full complement of cortical neurons is present by the 18th week of gestation, and by the ninth month the cortex comprises six distinguishable layers: The cerebral cortex is composed of two cell types: granular and agranular. Granular cortex is so-called because it contains a large percentage of granule cells, which receive afferent impulses. This type of cortex is located mainly in primary sensory regions, such as visual, auditory, and so-matosensory cortices. Granular cortex contains strongly developed layers II and IV (refers to cortical layers). Agranular cortex is so-called because it contains a high percentage of pyramidal cells (as compared with granule cells). It is located mainly in areas that give rise to efferent impulses, such as the motor cortex, and the frontal eye field, which gives rise to prominent corticobulbar and corticospinal projections. Agranular cortex is characterized by strongly developed pyramidal layers III and V and less well developed granular layers. Fig. 13.4 Cortical layers. See Fig. 13.5. In 1909, the German histologist Korbinian Brodmann distinguished over 50 different regions of the cerebral cortex. He based these divisions on variations in cytoar-chitectural features, such as relative thickness, density, size, shape, and arrangement of cortical neurons. Brod-mann believed that these variations in cytoarchitecture should correspond to distinct physiological functions. In fact, only a limited number of the Brodmann’s areas have been related to specific functions. Among the most important are the following: somatosensory cortex (areas 3, 1, and 2); primary motor and premotor cortex (areas 4 and 6, respectively); the frontal eye field (area 8); primary visual cortex (area 17); visual association cortex (areas 18 and 19); auditory cortex (areas 41 and 42); auditory association cortex (area 22, part of Wernicke’s language comprehension area); Broca’s speech area (area 44); sensory association cortex (areas 5 and 7); and prefrontal cortex (areas 9, 10, 11, and 12). Fig. 13.5 Brodmann’s areas. A variety of projection fibers originate in the cerebral cortex. Examples of these corticofugal projections include the corticostriatal and corticothalamic pathways; cortical projections to the amygdaloid body and the hippocampus; and corticospinal, corticoreticular, and cortico-pontine pathways. Many of the corticofugal projections, including the latter three named pathways, pass through the corona radiata and then the internal capsule. Other parts of the cortical projection system include the optic radiation, connecting the lateral geniculate body with the visual cortex, and the acoustic radiation, connecting the medial geniculate body with the auditory cortex. See Fig. 13.6. The association fibers constitute most of the cerebral white matter. They provide connections between both adjacent and remote areas of the cortex. Among the most important association fibers are the following. The arcuate fibers, also known as U-fibers, occupy the immediate subcortical white matter. They interconnect adjacent cortical fields. The superior longitudinal fasciculus connects the frontal lobes with portions of the parieto-occipital and temporal convolutions. It lies just dorsal to the level of the insula. The frontotemporal portion of this projection system is called the arcuate fasciculus and is believed to connect the temporal (Wernicke) and frontal (Broca) areas of speech. The temporo-occipital or inferior longitudinal fasciculus connects the temporal and occipital lobes; and the uncinate fasciculus connects the frontal lobe with the temporal pole. The cingulum is one of the association bundles of the limbic system. It connects the subcallosal area with the parahippocampal gyrus. Fig. 13.6 Association fibers. See Fig. 13.7. The commissural fibers cross the midline between the two cerebral hemispheres, providing interconnections between them. Important groups of commissural fibers include the following:
Major Sulci and Fissures
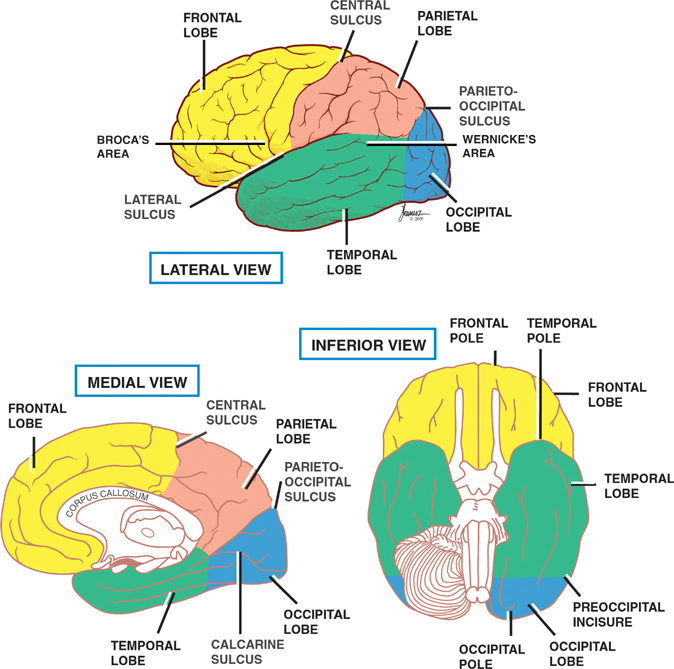
Gyri and Sulci
Frontal Lobe
Parietal Lobe
Temporal Lobe
Occipital Lobe
Medial Surface
Inferior Surface
Histology of the Cortex
Cortical Neurons
Cortical Layers
Granular and Agranular Cortex
Brodmann’s Map
Subcortical White Matter
Projection Fibers
Association Fibers
Commissural Fibers
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


