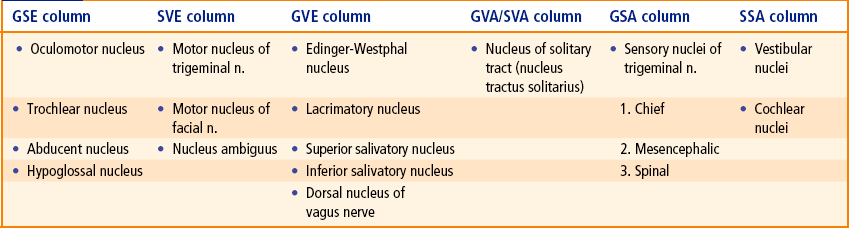
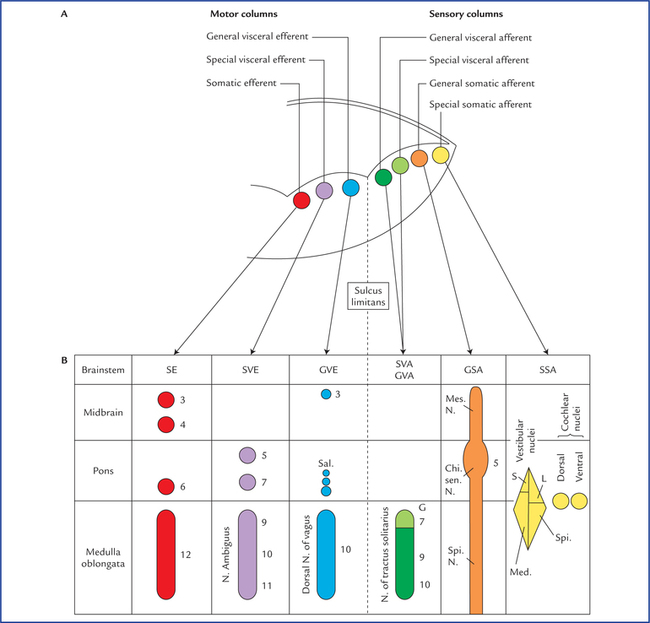
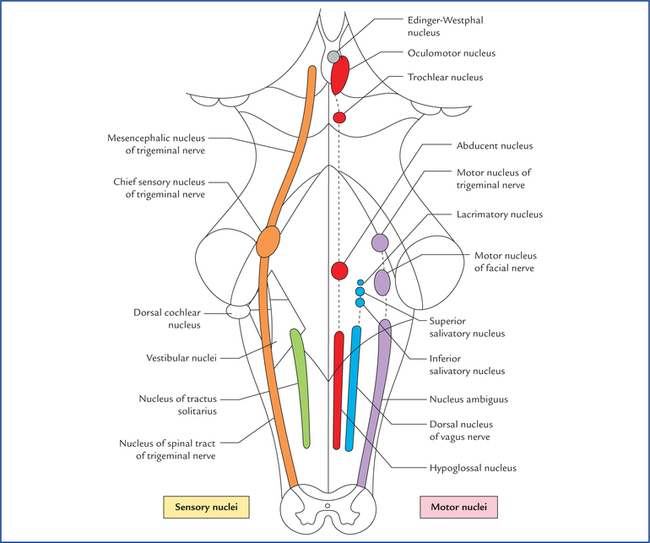
9 There are 12 pairs of cranial nerves, out of these two pairs arise from the forebrain and 10 pairs from the brainstem (see related text on page 47 and Figs 6.9, 8.1). The cranial nerves are designated by Roman numerals in order from before backwards in which they are attached on to the brain: The cranial nerves can be classified into the following three morphological groups: • Those supplying the muscles derived from cranial myotomes, viz. oculomotor (III), trochlear (IV), abducent (VI), and hypoglossal (XII) nerves. • Those supplying the muscles derived from branchial arches, viz. trigeminal (V), facial (VII), glossopharyngeal (IX), vagus (X), and accessory (XI) nerves. • Those associated with special sense organs, viz. olfactory (I), optic (II) and vestibulocochlear (VIII) nerves. 1. General somatic efferent column. 2. General visceral efferent column. 3. General visceral afferent column. Apart from this, an extra special somatic column appears in the most lateral part of the alar lamina to receive sensations of hearing and balance. Thus, there are seven functional columns in the brainstem (Fig. 9.1A), from medial to lateral side they are: Fig. 9.1 (A) Functional columns of grey matter in brainstem (of embryo). (B) Schematic diagram to show different cranial nerve nuclei in adult brain derived from seven functional columns of grey matter. (N = nucleus, Mes = mesencephalic, Spi = spinal, Chi Sen = chief sensory, S = superior, L = lateral, Med = medial, G = gustatory nucleus, EW = Edinger-Westphal, Sal = salivatory nuclei, 3 = oculomotor, 4 = trochlear, 5 = trigeminal, 6 = abducent, 7 = facial, 8 = glossopharyngeal, 9 = vagus.) As the development proceeds, these columns differentiate into one or more discrete cranial nerve nuclei. Nuclei derived from various functional columns are enumerated in Table 9.1 and shown in Figure 9.1B. The surface projection of these nuclei on the dorsal aspect of the brainstem is shown in Figure 9.2. Fig. 9.2 Surface projection of cranial nerve nuclei on the dorsal aspect of the brainstem. Motor nuclei are shown on the right side and sensory nuclei on the left side. • The general somatic efferent fibres arise from the nuclei of general somatic efferent column, and supply the striated muscles of the limbs and body wall developing from somites (‘soma’ = body wall). • The special visceral (branchial) efferent fibres arise from the nuclei of special visceral (branchial) efferent column and supply the striated muscles developing from branchial or pharyngeal arches, viz. muscles of facial expression, muscles of palate, pharynx and larynx. • The general visceral efferent fibres arise from the nuclei of general visceral efferent column and supply the glands and the smooth muscles of vessels and viscera. These fibres form the cranial outflow of the parasympathetic nervous system. • The general visceral afferent fibres carry general sensations (sense of distention and ischaemia) from the viscera, viz. lung, heart and upper part of GIT and associated glands to the general visceral afferent column. • The special visceral (branchial) afferent fibres carry special sensation of taste from tongue, etc. to the nuclei of special visceral afferent column (this is because the epithelium of tongue including taste buds develop from endoderm (visceral). • The general somatic afferent fibres carry general sensations (pain, touch and temperature from skin) and proprioceptive sensations (vibration, muscle and joint sense) to the nuclei of general somatic afferent column. • The special somatic afferent fibres carry special sensations of hearing and equilibrium to the nuclei of special somatic afferent column (this is because the organs concerned with these sensations develop from ectoderm of the body wall). General somatic efferent nuclei supply the striated muscles of somatic origin. Oculomotor nucleus is located in the central grey matter of midbrain, ventral to cerebral aqueduct at the level of superior colliculi (for details see page 85). Trochlear nucleus is located in the central grey matter of midbrain, ventral to cerebral aqueduct and close to midline at the level of inferior colliculi. It is just caudal to the oculomotor nucleus and its ventral aspect is closely related to medial longitudinal fasciculus. Fibres from each trochlear nucleus course dorsally and then medially around the central grey matter to reach the cranial end of superior medullary velum, wherein they decussate to emerge on the lateral side of frenulum veli on the dorsal aspect of the midbrain (Figs 8.3, 8.12), The trochlear nerve fibres have an unusual course and this is the only nerve which emerges from the dorsal aspect of brainstem. It has been suggested that this nerve originally supplied the muscles of pineal eye, which would account for its dorsal course. Hypoglossal nucleus is an elongated column consisting of motor cells like those of anterior horn cells of the spinal cord. It extends throughout the length of medulla oblongata in the paramedian plane. The upper part of nucleus lies deep to hypoglossal triangle in the floor of IVth ventricle. The medial longitudinal bundle lies immediately ventral to it. In the closed part (lower part) of medulla, the nucleus lies in the central grey matter ventral to the central canal. The fibres from the hypoglossal nucleus course ventrally on the lateral side of medial lemniscus and emerge on the ventral aspect of medulla as a series of about 12 rootlets in the sulcus between pyramid and olive to form hypoglossal nerve (Fig. 8.6). Special visceral efferent nuclei supply the striated muscles derived from branchial arches. Motor nucleus of trigeminal nerve is situated in the upper part of the pons, in its dorsal region, medial and just cranial to chief sensory nucleus of trigeminal nerve. Fibres from the motor nucleus constitute the motor root of trigeminal nerve which emerges on the ventral aspect of pons medial to the sensory root. The motor root crosses the superior border of petrous temporal bone and passes posterior to trigeminal ganglion. Then it passes through the foramen ovale and immediately joins the sensory root of the mandibular nerve (Figs 8.9, 9.4). Nucleus of facial nerve is situated in the lower part of the pons, in the ventrolateral part of its tegmentum, more or less in line with the motor nucleus of the trigeminal nerve. Its position is anterolateral and caudal to abducent nucleus, and medial to the nucleus of spinal tract of trigeminal nerve. The fibres arising from the nucleus pursue an aberrant course. First they course dorsomedially towards the floor of fourth ventricle to loop behind the motor nucleus of the abducent nerve. The loop (internal genu of facial nerve) elevates the floor of fourth ventricle and forms the facial colliculus, and then course ventrolaterally passing between the nucleus of their origin and nucleus of spinal tract of trigeminal nerve to emerge through the pontomedullary junction on the ventral aspect of the brainstem lateral to the emergence of the abducent nerve. The unusual course of motor fibres of facial nerve represents an example of neurobiotaxis (for details see page 80). Nucleus ambiguus is an elongated column of typical motor neurons, extending throughout the length of medulla. Nucleus ambiguus is so named because it is not clearly defined in sections of the medulla. It occupies a position dorsal to the inferior olivary nucleus and ventromedial to the nucleus of spinal tract of Vth nerve. Fibres from nucleus ambiguus are first directed dorsally and then turn sharply in the ventrolateral direction to mingle with other fibres of IXth, Xth, and XIth cranial nerves, which emerge from medulla along the posterolateral sulcus (Fig. 8.6). The fibres from nucleus ambiguus supply the muscles derived from third, fourth and sixth branchial arches. Edinger-Westphal nucleus is located in the upper part of the midbrain dorsal to the rostral two-thirds of the main oculomotor nucleus (Fig. 9.5). Preganglionic para-sympathetic fibres arising from this nucleus reach the ciliary ganglion by way of oculomotor nerve, where they terminate. The preganglionic fibres from superior salivatory nucleus pass to the submandibular ganglion through the nervus intermedius, geniculate ganglion, facial nerve and its chorda tympani branch (see Fig. 20.10). Preganglionic fibres from lacrimatory nucleus reach the pterygopalatine ganglion through nervus intermedius and greater superficial petrosal nerve (Fig. 20.10). Inferior salivatory nucleus is a small collection of nerve cells in the dorsolateral part of pons, just above its junction with the medulla. It lies immediately caudal to the superior salivatory nucleus and just above the upper end of the dorsal nucleus of the vagus nerve. Preganglionic fibres from this nucleus run in the glossopharyngeal nerve, enter its tympanic branch (nerve of Jacobson) and relay in the “otic ganglion” by way of tympanic plexus and lesser superficial petrosal nerve (Fig. 20.9). Motor nucleus of vagus is a long vertical column of cells extending throughout most of the length of medulla. Its upper end lies deep to the vagal triangle in the floor of the fourth ventricle. When traced downwards in the closed part of medulla, it occupies a position in the lateral part of central grey matter, dorsal to the hypoglossal nucleus (Fig. 8.5). This nucleus is the main source of parasympathetic fibres of vagus nerve. Nucleus of solitary tract is an elongated column of cells and is intimately related to a group of descending fibres which constitute the tractus solitarius. The upper part of the nucleus lies deep in the reticular formation, ventrolateral to the dorsal nucleus of vagus (Fig. 8.6). When traced downwards it lies in the dorsal part of central grey matter in the closed part of the medulla, dorsomedial to the dorsal nucleus of vagus (Fig. 8.5). The lower ends of the nuclei of two sides fuse to form the commissural nucleus of the vagus. The rostral portion of the nucleus is concerned with taste sensations and receives the special visceral afferent fibres from facial, glossopharyngeal and vagus nerves and is frequently referred to as gustatory nucleus. The nuclear terminations of VII, IX and X nerves are in rostrocaudal direction (Fig. 9.3). The caudal portion of the nucleus receives the general visceral sensations from pharynx (glossopharyngeal and vagus) and from oesophagus and abdominal part of alimentary canal up to right two-thirds of the transverse colon (vagus). It is presumed that axons from nucleus of tractus solitarius project to the thalamus and hypothalamus of the opposite side through the solitariothalamic and solitariohypothalamic tracts respectively. These tracts join the medial lemniscus of the opposite side on their way to thalamus and hypothalamus. The neurons from thalamus then project to the cerebral cortex (Fig. 9.3). Fig. 9.3 Connections of nucleus tractus solitarius. The inset on the right side shows that the lower ends of the nuclei of two sides fuse to form the commissural nucleus of the vagus nerve. Fig. 9.4 Schematic illustration to show connections of trigeminal nerve nuclei. M = motor nucleus. Inset on the right upper corner shows the subdivisions of spinal nucleus of trigeminal nerve and disposition of afferent fibres from its ophthalmic maxillary and mandibular divisions. (PR = pars rostralis, PI = pars inter-polaris, PC = pars caudalis, V1 = ophthalmic nerve, V2 = maxillary nerve, V3 = mandibular nerve, M = motor nucleus, SVE = special visceral efferent, GSA = general somatic afferent.) General somatic afferent nuclei include the three sensory nuclei of the trigeminal nerve. Chief sensory nucleus of trigeminal nerve lies in the dorsolateral region of the tegmentum of the upper part of the pons lateral to the motor nucleus (of trigeminal) and occupies an intermediate position between the mesencephalic nucleus above and the spinal nucleus below (Fig. 9.4). It is concerned only with the tactile sensibility. Spinal nucleus of trigeminal nerve extends caudally from chief sensory nucleus in the pons to the second cervical spinal segment and lies just medial to the spinal tract of trigeminal nerve. The spinal nucleus and tract of trigeminal nerve are chiefly concerned with the pain and temperature sensations. Based on the cytoarchitecture, the spinal nucleus is divided into three parts (or subnuclei): In craniocaudal direction these are: (a) pars rostralis, (b) pars interpolaris, and (c) pars caudalis (Fig. 9.4). The ascending fibres end in the chief sensory nucleus. The descending fibres from a large bundle of fibres called spinal tract of trigeminal nerve. Fibres of the spinal tract terminate in the subjacent spinal nucleus. The afferents from three trigeminal divisions rotate, so that the fibres of ophthalmic division terminate in the pars caudalis, the fibres of maxillary division in the pars interpolaris and the fibres of mandibular division in pars rostralis. Mesencephalic nucleus of trigeminal nerve extends from upper end of chief sensory nucleus in the pons to the midbrain where it lies in the central grey matter lateral to the cerebral aqueduct. Because it extends rostrally into the midbrain, it is called mesencephalic nucleus. Like dorsal root ganglia of spinal cord it is made up of pseudounipolar cells (1st order sensory neurons) and appears to have similar functions (the mesencephalic nucleus is unique in the sense that it is the only site in CNS which contains the cell bodies of first order sensory neurons). Peripheral processes of these cells carry proprioceptive impulses from the muscles of mastication, temporomandibular joint, teeth and possibly also from the extrinsic muscles of tongue. Central processes terminate in the motor nuclei of trigeminal nerve of the both sides. These connections establishes the stretch reflex originating in the neuromuscular spindles in masticatory muscles, together with a reflex for control of the force and accuracy of bite (Barr, N.L., 1972). These reflexes prevent the tongue from being bitten during chewing. Other central processes relay in the cells of reticular formation. From which fibres arise and run through dorsal trigeminal lemniscus to relay in the ventral posteromedial nucleus (VPM) of the thalamus (Fig. 9.4). The cochlear nuclei contain the cell bodies of the second order sensory neurons in the auditory pathway. Their connections are described in Chapter 18. Vestibular nuclei are situated partly in the medulla and partly in the pons, immediately beneath the lateral part of the floor of the fourth ventricle called vestibular area. On the basis of cytoarchitecture and afferent and efferent connections, four distinct vestibular nuclei are recognised, viz. (a) inferior or spinal vestibular nucleus, (b) lateral vestibular nucleus (also called Dieter’s nucleus), (c) superior vestibular nucleus, and (d) medial vestibular nucleus (Fig. 8.15). • Inferior vestibular nucleus lies in the medulla, just medial to the inferior cerebellar peduncle. It is continuous rostrally with the lateral vestibular nucleus and related medially to the medial vestibular nucleus. It extends from the cranial end of nucleus gracilis to the pontomedullary junction. • Lateral vestibular nucleus lies immediately cranial to inferior vestibular nucleus and extends upwards in the pons almost to the level of nucleus of abducent nerve. It is composed of large multipolar cells resembling typical motor neurons. The cells of this nucleus give origin to the fibres of lateral vestibulospinal tract (Fig. 8.15). • Superior vestibular nucleus is smaller in size and located entirely within the pons above the medial and lateral vestibular nuclei. • Medial vestibular nucleus extends from medulla at the level of olive to the lower part of the pons. It is bounded laterally and rostrally by the other three vestibular nuclei. Its medial border is near the midline of the brainstem. The caudal end of this nucleus is near the caudal limit of the fourth ventricle.
Nuclei, Functional Components and Distribution of Cranial Nerves
Morphological Classification of Cranial Nerves
Functional Columns and Nuclei of Cranial Nerves
Development of Functional Columns and Nuclei of Cranial Nerves

General Somatic Efferent Nuclei
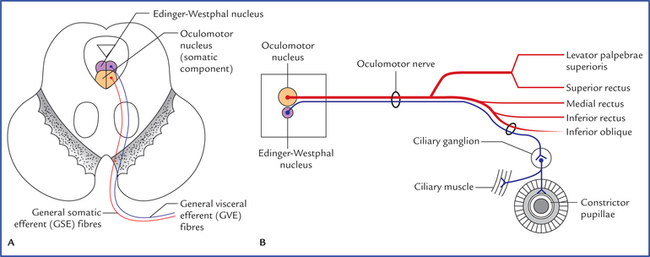
Oculomotor nucleus
Trochlear nucleus
Hypoglossal nucleus
Special Visceral (Branchial) Efferent Nuclei
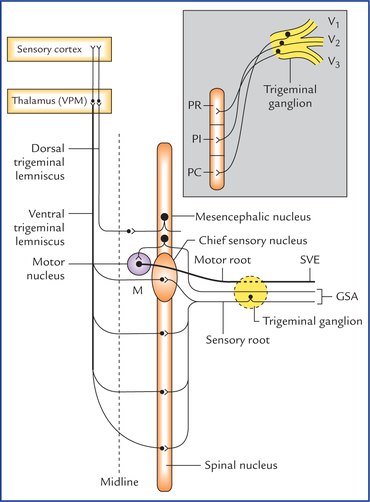
Motor nucleus of trigeminal nerve
Nucleus of facial nerve
Nucleus ambiguus
General Visceral Efferent Nuclei
Edinger-Westphal nucleus (visceral oculomotor nucleus)
Superior salivatory and lacrimatory nuclei
Inferior salivatory nucleus
Dorsal nucleus of vagus (also called motor nucleus of vagus)
General and Special Visceral Afferent Nuclei
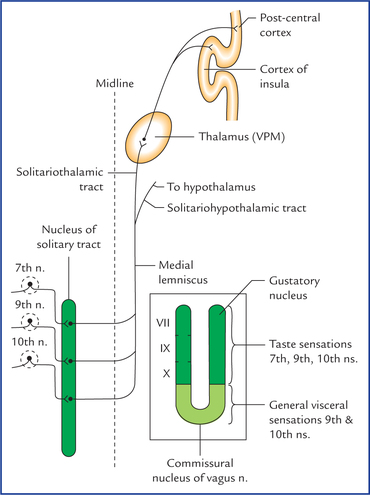
Nucleus of solitary tract (Fig. 9.3)
General Somatic Afferent Nuclei
Chief sensory nucleus of trigeminal nerve
Spinal nucleus of trigeminal nerve
Mesencephalic nucleus of trigeminal nerve
Special Somatic Afferent Nuclei
Cochlear nuclei
Vestibular nuclei
![]()
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


Nuclei