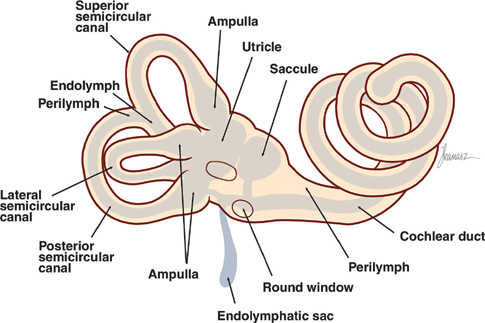
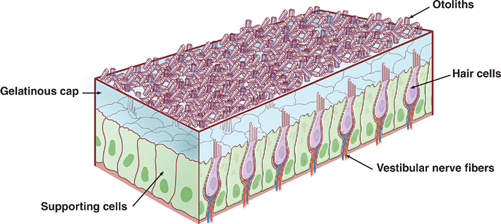
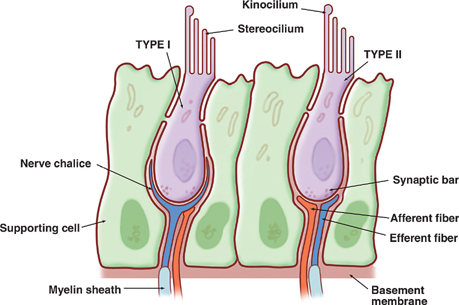
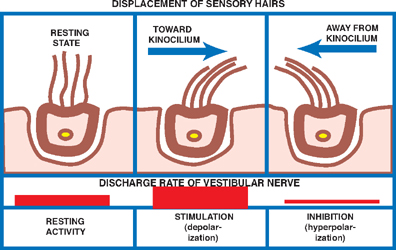
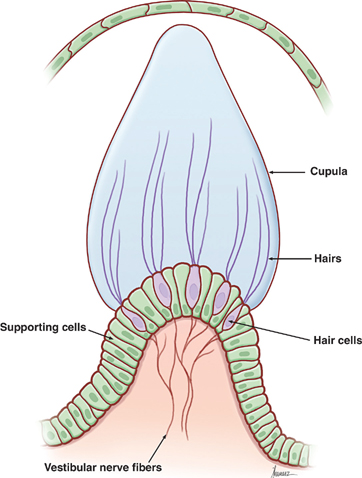
17 Vestibular System Dizziness is one of the most common complaints for which patients seek the attention of a neurologist. Often the underlying disorder causing patients to experience dizziness is disequilibrium, which represents malfunction in one of three separate but communicating systems: the visual system, the proprioceptive system, or the vestibular system. As will be seen, the diagnosis of disequilibrium as a disturbance of the vestibular system is based on a constellation of signs and symptoms elicited from the patient at bedside. Much is understood about the anatomy and physiology of the vestibular system, the basic functions of which include the coordination of motor control, posture, equilibrium, and eye movements. This chapter begins with a description of the functional anatomy of the peripheral vestibular system. Next, central connections with the spinal cord, cerebellum, and oculomotor system are discussed, followed by a practical approach to the dizzy patient. See Fig. 17.1. The peripheral vestibular system detects movement and position of the head in space. The inner ear, or labyrinth, is composed of two parts: a bony labyrinth formed by cavernous openings in the petrous portion of the temporal bone, and a membranous labyrinth formed by a simple epithelial membrane. The membranous labyrinth, which is filled with a fluid called endolymph, lines the contours of the bony labyrinth from which it is separated by a fluid called perilymph. Peripheral receptors of the vestibular and auditory systems are contained within the membranous labyrinth. Two sets of structures comprise the vestibular portion of the labyrinth: (1) the utricle and saccule and (2) the semicircular canals. Fig. 17.1 Peripheral vestibular system. See Figs. 17.2, 17.3, 17.4, and 17.5. The utricle and saccule detect linear acceleration and the position of the head in space. These structures are dilatations of the membranous labyrinth, separated (by perilymph) from the bony labyrinth. They are filled with endolymph and lined by simple cuboidal epithelium except at receptor-rich regions called maculae. Utricular and saccular maculae are histologically identical. They contain the following four elements: (1) supporting cells, which consist of simple columnar epithelial cells that are continuous with the simple cuboidal epithelial cells that line the utricle and saccule; (2) hair cells, which are specialized receptor cells intercalated between the supporting cells; (3) an otolithic membrane, which is a gelatinous mass embedded with crystals of calcium carbonate (otoliths) that covers the hair cells; and (4) dendrites of cells in the vestibular ganglion, which carry afferent impulses from the hair cells to the vestibular nuclei of the brainstem. The “hairs” of a macular hair cell are microvilli. Each hair cell contains ~40 to 80 microvilli together with a single kinocilium that arises from a centriole. One kino-cilium is located in the periphery of each hair cell, thus polarizing the hair cell in the direction of the kinocilium. The kinocilia of the maculae of the utricle and saccule are arranged in such a way as to polarize the maculae in relation to an imaginary curved line called the striola. Utricular maculae are polarized toward the striola, saccular maculae are polarized away from the striola. The functional significance of this arrangement is as follows. The weight of the otolithic membrane exerts a gravitational pull on hair cells. The orientation of the hair cells relative to this gravitational force determines the direction of their displacement. For example, when the head is held in the horizontal plane, the vertical gravity vector is perpendicular to the hair cell, and no displacement occurs. When the head is tilted, say, to the left, hair cells follow, and the gravitational force of the oto-liths, now at an angle to the cells, deflects them further in this direction. Fig. 17.2 Histology of the utricle and saccule. Fig. 17.3 Hair cell histology. Displacement of hair cells in the direction opposite to the axis of polarization hyperpolarizes the cells and initiates inhibitory impulses. Thus, in response to a tilt of the head, certain hair cells depolarize, whereas others hyperpolarize—depending on their axis of polarization (i.e., the relation of their kino-cilium to the striola). Because of their multiple axes of polarization (a product of the curved striola), the utricle and saccule detect changes of head position in any direction in the vertical (saccule) and horizontal (utricle) planes. The overall pattern of macular activity—of excitation and inhibition in both the utricle and the saccule—is finally relayed to the vestibular nuclei, where it signals the position of the head in space. Although they predominantly function as sensors of static position, the utricle and saccule detect linear acceleration as well. Fig. 17.4 Polarization of utricular and saccular maculae. Fig. 17.5 Sensory hairs. See Figs. 17.1, 17.6, 17.7, and 17.8. The three endolymph-containing semicircular ducts detect angular acceleration of the head in space are formed by the membranous labyrinth, in direct continuation with the utricle. Because the ducts lie in planes oriented at right angles to one another, they provide information about the angular acceleration of the head in any direction, though they lack the ability of the utricle and saccule to detect its static position. One end of each duct enlarges at its point of attachment to the utricle. These enlargements, or ampullae, are the functional counterparts of the maculae of the utricle and saccule. They contain a thickened sensory epithelium called the ampullary crest, which is composed of columnar epithelial supporting cells with interspersed kinocilia-containing hair cells. These hair cells are embedded in a gelatinous mass termed the cupula. They fill the space between the crest and the roof the ampulla. (The cupula is the equivalent of the otolithic membrane but lacks its high-specificgravity calcium carbonate crystals.) Angular acceleration of the head causes displacement of endolymphatic fluid and movement of the cupula, which stimulates the hair cells. The kinocilia of the horizontal ducts are polarized in the direction of the utricle. Bending hair cells of these ducts in the direction of the utricle thus results in excitatory output. Conversely, the kinocilia of the vertical ducts are polarized in the direction opposite the utricle. Bending hair cells in these ducts in the direction of the utricle thus results in inhibitory output. In a given plane, the axis of polarization of the ampullae of a pair of ducts, one in the right ear and one in the left, are mirror images of one another. Deflection of their cupulae by the clockwise or counterclockwise flow of endolymphatic fluid excites the duct whose hair cells are deflected toward the axis of polarization, while inhibiting the duct whose hair cells are deflected opposite their axis of polarization. The combined effect of excitatory and inhibitory input from paired semicircular ducts on opposite sides of the head apprises the brain of the head’s rotary movements.
Functional Anatomy
The Peripheral Vestibular System
The Utricle and Saccule

The Semicircular Ducts
Related posts:






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


