and Robert E. Schmidt2
(1)
Sunnybrook and St Michael’s Hospitals, University of Toronto, Toronto, ON, Canada
(2)
Division of Neuropathology Department of Pathology, Washington University School of Medicine, St. Louis, MO, USA
6.1 Normal Structure and Function
6.1.1 Vascular Anatomy
Nutrient arteries arise from major vessels and penetrate the nerve trunk roughly perpendicular to its axis. These vessels are highly variable in caliber (up to 1 mm) and in location, both between and within (from side to side) individuals. The median and ulnar nerves, for example, have 1–11 (mean 3.24) and 2–19 (mean 7.75) such nutrient arteries in their forearm segments, respectively (Sunderland 1978). After entering the nerve trunk, the nutrient vessels divide into longitudinal ascending and descending branches and supply several arrays of vessels which run along the nerve and interconnect via oblique or laterally directed channels.
Lundborg (Lundborg 1970; Lundborg and Branemark 1968) has divided the neural vascular system into extrinsic and intrinsic parts. The extrinsic system consists of the “… regional nutrient arteries which supply the Intraneural vascular bed at varying intervals.” The intrinsic system is contained within the nerve and consists of longitudinal arrays of vessels running along the nerve, with abundant anastomosing channels between them. This system includes both epineurial and endoneurial vessels. More recently, some authors have used the term “intrinsic” to mean the intrafascicular vascular plexus (McManis et al. 1993; Olsson 1972).
The epineurial (extrafascicular) intrinsic vessels include arterioles which run longitudinally at the most superficial aspects of the epineurium. These arterioles are sometimes visible on the surface of the nerve. The epineurial intrinsic vessels (Fig. 6.1a) also include an array of arterioles deeper within the nerve, located between individual fascicles and vasculature within perineurial septa (Fig. 6.1b). A cross section of the nerve trunk may show several arterioles, but one usually dominates (arrowhead, Fig. 6.1a). The lack of an elastic lamina distinguishes venules which outnumber arterioles (Figs. 6.1a and 6.2a, b).
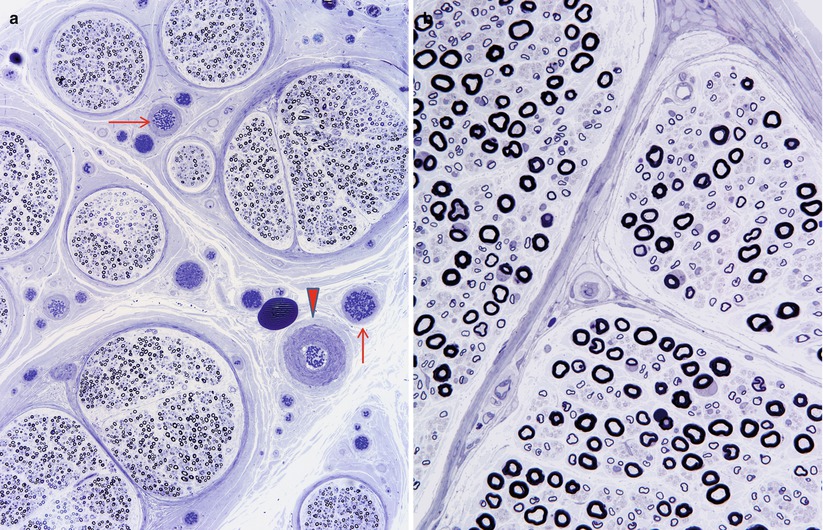
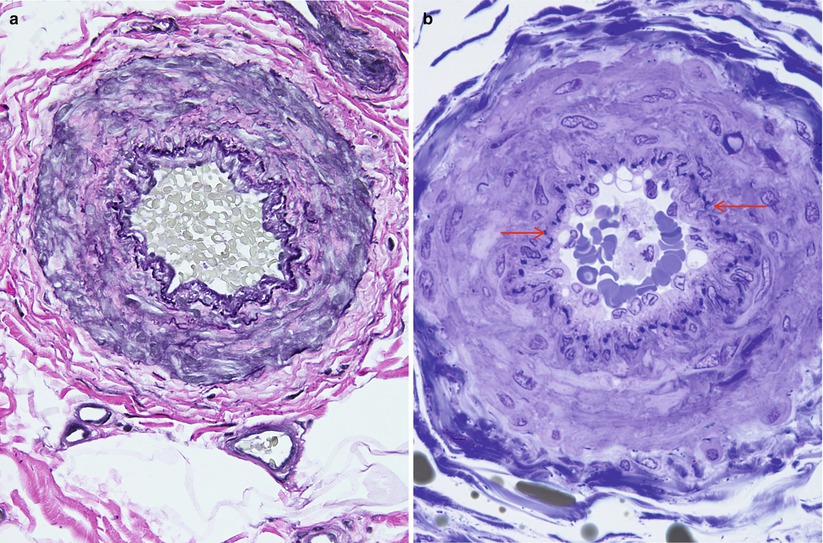
Fig. 6.1
One arteriole (arrowhead) and several venules (arrows) are shown in the epineurium (a). In (b) note vessels within perineurial septum (1 μm thick toluidine blue-stained plastic section, 1,000×)
Fig. 6.2
(a) Epineurial arteriole is stained with Verhoeff–Van Gieson elastica. In semithin sections the internal elastic lamina (arrow) is less conspicuous (b) (a: paraffin, b: 1 μm thick toluidine blue-stained plastic section)
An endoneurial capillary system runs longitudinally with numerous oblique and transverse anastomoses among vessels (Figs. 6.1a and 6.3). The literature is somewhat unclear about whether arterioles are found in the endoneurium. Bell and Weddell (1984) described endoneurial arterioles having one or two layers of smooth muscle cells and an incomplete internal elastic lamina, but Sunderland (1978) has stated that arterioles are not seen in the endoneurium. This disagreement may simply be a nosologic issue, depending on the definition of endoneurium and of arterioles. Beggs et al. (1991) noted that true arterioles, when found inside a fascicle, are confined to intrafascicular perineurial septa and are not in the endoneurium, strictly speaking (Fig. 6.4). Moreover, the presence of arterioles may vary depending on the nerve trunk and species under study (Beggs et al. 1991), for the studies of Bell and Weddell (1984) were performed on sciatic nerves of several species including humans. We have never found an intrafascicular arteriole outside a perineurial septum.
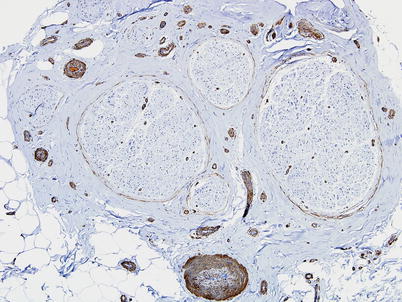
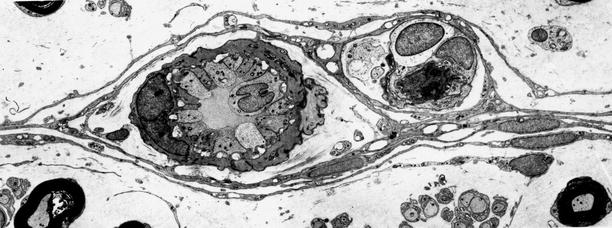
Fig. 6.3
Smooth muscle actin immunostaining highlights the epineurial and endoneurial microvasculature (paraffin, 100×)
Fig. 6.4
Terminal arteriole in perineurial partition (2,070×)
The endoneurial capillary density has been measured at 60–100 per mm2 in human sural nerve, decreasing somewhat with age (Dyck et al. 1985; Giannini and Dyck 1993). These endoneurial microvessels differ from those of other organs (Bell and Weddell 1984). The presence of strong alkaline phosphatase activity suggests their essential capillary nature. However, most vessels are associated with a periendothelial cell with cytoplasm extending over 50 % or more of the circumference and are thus reminiscent of postcapillary venules. Furthermore, endoneurial microvessels are larger than those of other tissues, with a mean diameter of 9 μm vs. 5.2 μm in muscle, for example (Bell and Weddell 1984). Giannini and Dyck (1993) found endoneurial microvessel diameters ranging from 5 to 22 μm. Endoneurial capillaries are lined by a single layer of thin endothelial cells, about five per vessel cross section, linked by tight junctions at points of contact (Giannini and Dyck 1993) (Figs. 6.5, and 6.6a). Junctions are associated with a large variety of molecules (claudins 1,2,5,12,19; zona occludens (ZO) 1,2; and other adhesion molecules, Ubogu 2013). Capillary endothelial cells contain Weibel–Palade bodies (Fig. 6.6a) which are thought to produce factor VIII-related antigen (Rondaij et al. 2006). Endothelial cells may also express numerous chemokines, cytokines, and growth factors, which may participate in neuropathy. Normal endothelial cells of the epineurium and perineurium may demonstrate fenestrae, but those of the endoneurium do not. Immediately around the capillary is a condensation of the collagen found throughout the endoneurium. However, in contrast to vascular structures in the central nervous system, there is a generous extracellular space around vessels, and no “glial” cells invest the walls of the blood vessel.
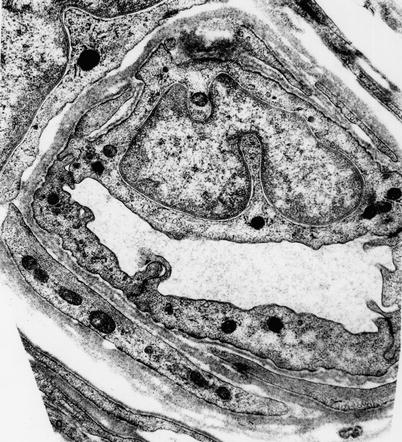
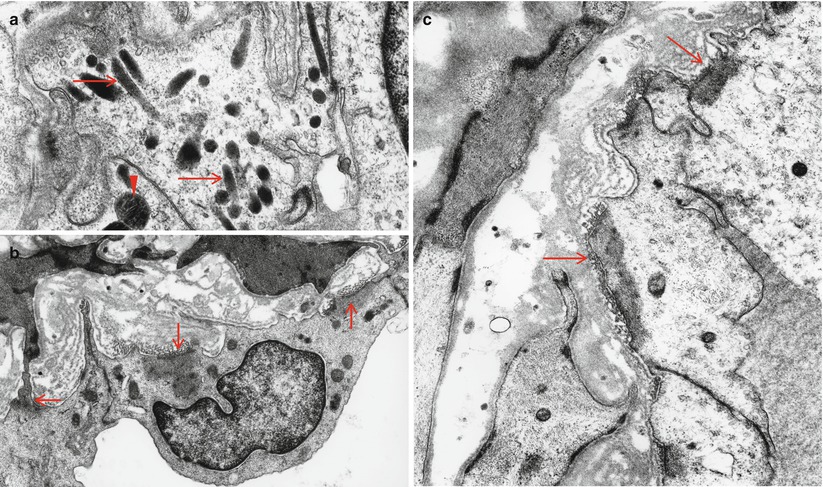
Fig. 6.5
Typical endoneurial microvessel displays intense pinocytotic activity, tight junctions, and pericytic component (1,3794×)
Fig. 6.6
(a) Large number of Weibel–Palade bodies (arrows) in endoneurial capillary endothelium. This patient suffered from Fabry disease: a single lipid inclusion is also seen (arrowhead). In (b, c) a complex of basal lamina condensation and anchoring filaments opposite focal filamentous accumulations (arrows) identifies these vessels as lymphatics (a 37,180×; b 19,760×)
Transperineurial arterioles connect the epineurial and endoneurial vessel arrays. Beggs and colleagues (1991) have defined transperineurial arterioles as vascular segments exhibiting a continuous smooth muscle coat and confined to the perineurial compartment, including all arterioles in the perineurium proper and arterioles within perineurial septa in the endoneurium (Fig. 6.5). These arterioles, which measure 10–25 μm in diameter, travel an oblique or transverse route through the perineurium (Beggs et al. 1991). The presence of axon terminals on the smooth muscle cells of these vessels suggests a potential for neurogenic regulation of their caliber (Beggs et al. 1991).
An incomplete smooth muscle cell layer and endothelial cells that are somewhat thinner than those of arterioles identified endoneurial venules (Bell and Weddell 1984). The endoneurial vessels drain into epineurial venules, which outnumber epineurial arterioles, and ultimately flow into large vessels which exit the nerve trunk along with the nutrient arteries.
A lymphatic drainage system has been described in the epineurium, but not in the endoneurium (Sunderland 1978). We have found that some large vessels in the epineurium demonstrate focal aggregations of cytoplasmic filaments in endothelial cells adjacent to focal condensation of basal lamina (Fig. 6.6b, c). The frequency with which these complexes are seen suggests that they are lymphatic vessels and a normal finding.
The endoneurium and subperineurial interstitial spaces are contiguous along the length of the nerve; consequently, material injected into the endoneurial compartment can be detected at considerable lengths up and down the nerve trunk, although it does not spread into the epineurium. Indeed, the endoneurial interstitial compartment is contiguous with the subarachnoid space (Olsson 1990).
6.1.2 Resistance of Peripheral Nerve to Ischemia
Overall then, there are four longitudinal arrays of vessels; superficial epineurial, deep epineurial, perineurial, and endoneurial, each of progressively lesser caliber. Typical lumina are 75–250 μm for epineurial arterioles, 10–25 μm for transperineurial arterioles, and 5–20 μm for endoneurial capillaries (Beggs et al. 1991; Dyck et al. 1972, 1985, 1987; Giannini and Dyck 1993). Numerous interconnections exist within and between the intra- and extrafascicular systems, providing the nerve with a high resistance to focal ischemia.
In vivo studies using rabbit tibial nerve demonstrate that occlusion of all nutrient vessels over a several centimeter segment does not cause visible change in blood flow, nor does the additional stripping of all epineurial vessels (Lundborg and Branemark 1968). Presumably, longitudinal flow within the intrafascicular circulation is able to compensate. Conversely, sectioning a nerve 3 cm above and below the observation point, thus eliminating the longitudinal intrafascicular blood flow, does not result in visible ischemia. Hypothetically, blood flow is maintained because the nutrient arteries feed the epineurial vessels, which interconnect readily with the intrafascicular capillaries. Indeed, two nutrient arteries are sufficient to maintain a nerve segment in the absence of the longitudinal intrafascicular circulation (Lundborg and Branemark 1968). Recent studies of peripheral nerve ischemia verify that endoneurial blood flow is dependent on both regional anastomoses with epineurial vessels through transperineurial arterioles and on longitudinal intrafascicular flow (Myers et al. 1991).
Peripheral nerve vasculature demonstrates an impressive functional reserve capacity, with numerous channels capable of opening in response to only a slight increase in nerve temperature or the sectioning of the other vessels (Lundborg 1970). Sympathetic activity also modulates the lumen of muscular vessels (epineurial, perineurial, transperineurial) and hence neural blood flow (Beggs et al. 1991; Kihara and Low 1990; Lundborg 1970).
6.1.3 The Blood–Nerve Barrier
The endoneurial compartment has a specialized ionic and macromolecular milieu (Ubogu 2013) thought to be important in nerve function, especially impulse propagation (Olsson 1972). Endoneurial fluid is under hydrostatic pressure greater than that of epineurial fluid, is hypertonic to plasma, and has less protein than plasma (Low et al. 1977; Myers et al. 1983). Maintenance of this unique composition requires isolation of the endoneurium from the vascular compartment and from the epineurium. Such isolation is a function of the blood–nerve barrier (BNB) and the perineurial barrier (PB) reviewed by Olsson (1990) and Ubogu (2013).
Early studies showed that, when injected into the experimental animal’s circulation, various markers would penetrate minimally, or not at all, into the endoneurial compartment (Olsson 1972, 1990). Microscopic studies suggested that the site at which passage was blocked or markedly slowed was the innermost layer of the perineurium and the normally non-fenestrated endothelial cells of the endoneurium; the electron-dense tight junctions found at both of these sites form the anatomical basis of the PB and BNB, respectively. Vessels within the epineurium and perineurium contain numerous fenestrations and lack tight junctions. Studies using a mast cell degranulation promoting agent given intravascularly to experimental animals demonstrated that this structural barrier was also a functional barrier. Mast cell contents were liberated in the epineurium but not in the endoneurium (Olsson 1972). Substantial interspecies variation exists in the BNB (Olsson 1972). Consequently, the extent to which one can apply data derived from animals to human tissues in health and disease is uncertain.
Albumin can enter the endoneurium (Poduslo 1993), and immunohistochemical studies on normal nerve biopsy specimens suggest that IgG and to a lesser extent IgA also can penetrate this compartment, while IgM and complement (C3) probably cannot enter in significant amounts (Liebert et al. 1985; Schenone et al. 1988; Takatsu et al. 1985). These observations indicate that the BNB is a relative barrier, meaning that one macromolecule (e.g., IgA) might have a much slower diffusion or transport rate into the endoneurial compartment than another (e.g., IgG), but given sufficient time and a large enough concentration gradient between the two compartments, a certain amount of both macromolecules will accumulate (Mata et al. 1987). Accumulation of macromolecules in the subperineurial area is a common finding in normal and abnormal nerves (Graham and Johnson 1985; Liebert et al. 1985; Schenone et al. 1988; Van Lis and Jennekens 1977) and should not be ascribed undue significance.
This blood–nerve barrier is leaky in the spinal nerve roots and in the dorsal root and autonomic ganglia allowing access to exogenous tracers, some toxins, and plasma proteins. This has been attributed to a lack of tight junctions between the capillary endothelial cells of the endoneurial compartment at these sites.
6.2 Pathological Alterations
6.2.1 Alteration in the Blood–Nerve Barrier in Neuropathy
Many peripheral neuropathies alter the BNB. Diabetes, Guillain–Barré syndrome, paraproteinemic neuropathy, leprosy, and lead exposure are examples of extensively studied peripheral nerve diseases where careful observation of the BNB indicates that it may be abnormal at times and that it potentially plays an important part in the pathogenesis of the neuropathy (see respective chapters for details). Whether such alterations in the BNB are primary or secondary to the neuropathy is a central question that has not been resolved in most cases (vide infra). In theory at least, an altered BNB can permit toxic substances to enter nerve and may cause increased endoneurial pressure with resultant reduced blood and nutrient flow, resulting in changes in the composition of the endoneurial milieu. These changes in turn may induce pathological processes such as fibrosis (Olsson 1990). An influx of vascular fluid into the endoneurium may result in alterations of the electrolyte concentration which permit the normal electrical function of peripheral nerve axons (Myers et al. 1983).
Immunohistological techniques permit study of alterations in the BNB in various human neuropathies (Liebert et al. 1985; Neuen et al. 1987; Van Lis and Jennekens 1977). IgM, ordinarily excluded from the endoneurium, frequently appears within it in inflammatory and vasculitic neuropathies. On the other hand, endoneurial IgM does not typically appear in metabolic, hereditary, or toxic neuropathies. An increase in the endoneurial content of smaller macromolecules such as IgG and albumin occurs in some noninflammatory neuropathies, representing perhaps a more subtle change in the BNB than the one which permits IgM to enter. In one study, the severity of disruption in the BNB correlated better with the presence of inflammation than with the severity of myelin or axon damage (Neuen et al. 1987).
An accumulation of macromolecules beneath the inner perineurial layer is commonly seen, perhaps due to entry via the endoneurial capillaries but blockage at the perineurium (Van Lis and Jennekens 1977). This accumulation of macromolecules seems to be particularly prominent in hypertrophic neuropathies (Van Lis and Jennekens 1977) and may correlate with the frequent finding of subperineurial edema in this setting.
Endoneurial vessel fenestration is always abnormal (Fig. 6.7). While more commonly associated with leprosy (Boddingius 1977, 1984), CIDP (Johnson 1977), and paraproteinemic neuropathy (Lach et al. 1993), this alteration is also found in diabetic neuropathy (Powell et al. 1985) and Wallerian degeneration (Ohara and Ikuta 1985). In addition to these settings, we have also found endoneurial fenestration in chronic axonal neuropathies of unknown etiology.
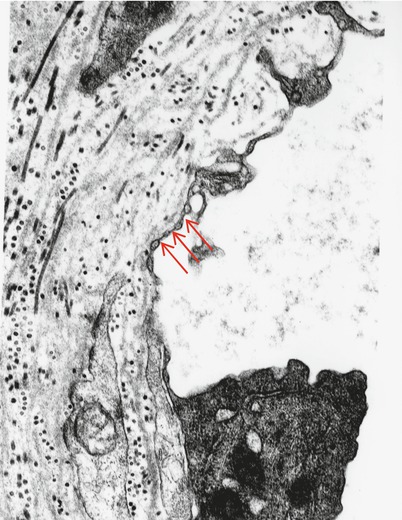
Fig. 6.7
Fenestration of endoneurial endothelium is shown (arrows) in a patient with CIDP (25,536×)
6.2.2 Endoneurial “Edema”
An increase in the amorphous or clear interstitial space of the endoneurium and subperineurial area is a common and nonspecific light microscopic finding. However, this finding is most closely associated with the hypertrophic neuropathies, familial or acquired (Behse et al. 1974; Matthews et al. 1970) (Fig. 6.8). The increased separation between nerve fibers and widening of the subperineurial space has been referred to as “endoneurial edema” and “subperineurial edema,” respectively. This “edema” is lightly eosinophilic, stains weakly with Alcian blue or toluidine blue, and does not stain with PAS or Congo red (Asbury et al. 1971). Electron microscopy shows no specific features except for dispersal of normal elements, although at times a fine granular substance accumulates. Possibly this substance represents “acid mucopolysaccharide” material in keeping with the histostaining properties described above. Such granular material has not been a significant finding in biopsies we have examined; perhaps the granular appearance is an artifact related to fixation or preparation.
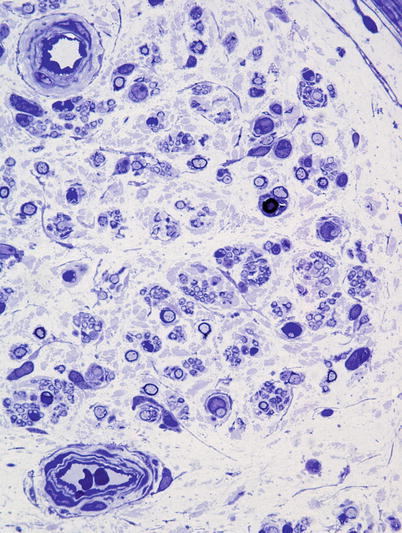
Fig. 6.8
Dispersion of endoneurial contents by increased intercellular matrix in CIDP (1 μm thick toluidine blue-stained plastic section, 1,000×)
The tendency of macromolecules to accumulate in the subperineurial area in both normal and pathologically altered nerves (Van Lis and Jennekens 1977) may relate to the frequent observation of subperineurial “edema.” Watanabe and Ohnishi (1979) demonstrated a significant enlargement of the subperineurial space in idiopathic polyradiculoneuropathy and thiamine deficiency-associated neuropathy. No correlation was found between prominence of subperineurial “edema” and severity of fiber damage or duration of illness (Watanabe and Ohnishi 1979). Such subperineurial space enlargement has also been described in a variety of inflammatory, toxic, metabolic, and inherited neuropathies.
Regions of endoneurial “edema” may be related to Renaut bodies. A fibrillar precursor of elastic fibers, oxytalan, is frequently seen at sites of “edema” and in Renaut bodies. Fibroblasts with giant vacuoles are also often detected in both of these locations. Enlarged subperineurial regions devoid of axons and containing few or no cellular elements are often described as “subperineurial edema,” but when fibroblastic configurations typical of Renaut bodies (Figs. 2.13c, d and 2.15a, b) are seen at this site, the distinction between “edema” and Renaut bodies becomes blurred. Certain ill-defined environmental conditions induce structural and functional changes in fibroblasts that result in the giant vacuolated appearance and in the production of acid mucopolysaccharide material and oxytalan (i.e., “edema”). In some instances, this process may proceed further to the formation of Renaut bodies. Indeed, precisely such a sequence of events has been reproduced experimentally in chronic nerve compression (Ortman et al. 1983).
While nerve “edema” may reflect deposition of acid mucopolysaccharides or oxytalan, or may represent a fixation artifact, it is also possible that this appearance may result from an osmotic increase in interstitial fluid. A balance of hydrostatic and osmotic pressures regulates endoneurial water content (Olsson 1990) and an increase in the macromolecular content of the interstitial compartment will cause an influx of fluid. This increase may result from pathological vascular permeability permitting an influx of albumin and other macromolecules, but can also theoretically occur without alteration in the BNB. For example, an influx of fluid could occur as a consequence of the release of macromolecules by endoneurial cells or the entrapment of macromolecules within the endoneurium by biochemical modification (Olsson 1990).
An increase in endoneurial interstitium is not always due to “edema.” We have seen cases of IgM paraproteinemic neuropathy where massive buildup of IgM in the endoneurium was present and caused the neuropathy. In this setting the acellular spaces were PAS positive.
6.2.3 Significance of Alterations in the Blood–Nerve Barrier
A central question is whether the increase in BNB permeability seen in neuropathy is secondary to axon and/or myelin damage or whether the permeability change is primary, with resultant nerve damage. Studies of Wallerian degeneration and toxic neuropathy demonstrate that the BNB normally loses its integrity during the early phases of axonal degeneration or demyelination and gradually recovers over several months, usually in parallel with return of axons and their myelin sheaths (Bouldin et al. 1991; Ohara and Ikuta 1985; Seitz et al. 1989). Latker et al. (1991) showed that the BNB recovered in the absence of nerve fiber regeneration, while the PB did not. In a lead neuropathy model, permeability and fluid contents changed only after substantial quantities of lead had entered the endoneurial compartment (Poduslo 1993). Although such data suggests the primacy of axon and Schwann cell changes, alterations in vascular permeability and endoneurial fluid composition have frequently been postulated to be contributors to the severity of neuropathy, in diseases ranging from diabetes and inflammatory neuropathies to leprosy and amyloidosis (Boddingius 1977; Brosnan et al. 1990; Hahn et al. 1985; Koski 1992; Poduslo 1993). Computer models and in vivo experimental work suggest that transperineurial vessels, especially venules, will become distorted and compressed when endoneurial pressure is increased, possibly resulting in reduced blood flow and injury to neural elements (Kalichman and Myers 1991; Myers et al. 1986, 1991).
Related posts:






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


