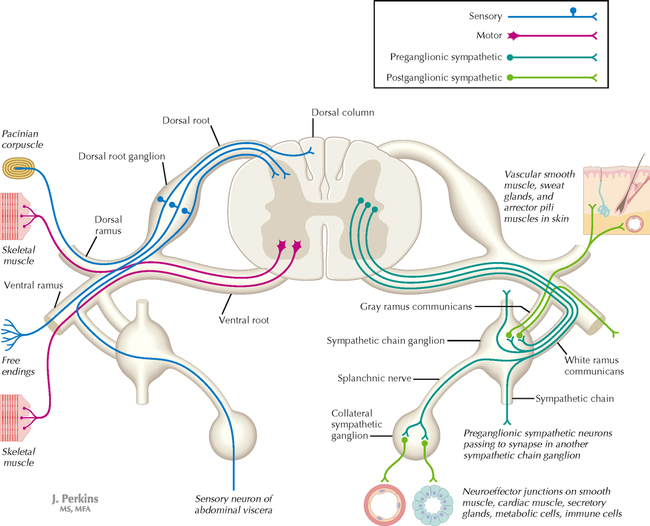
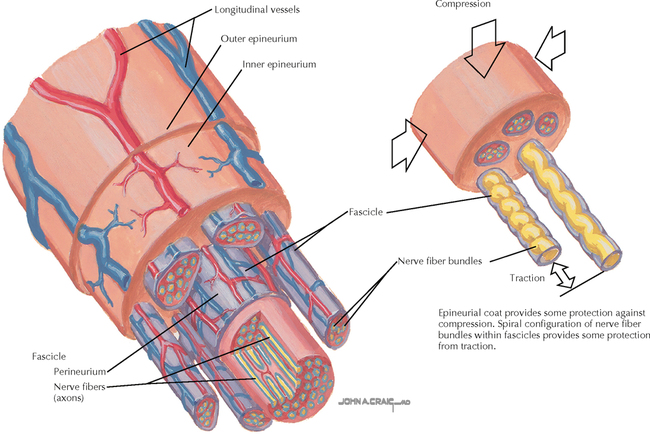
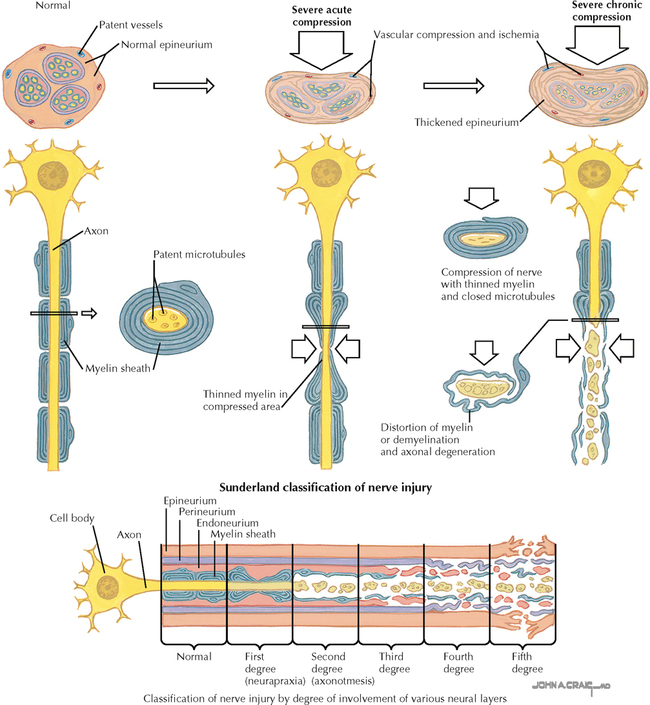
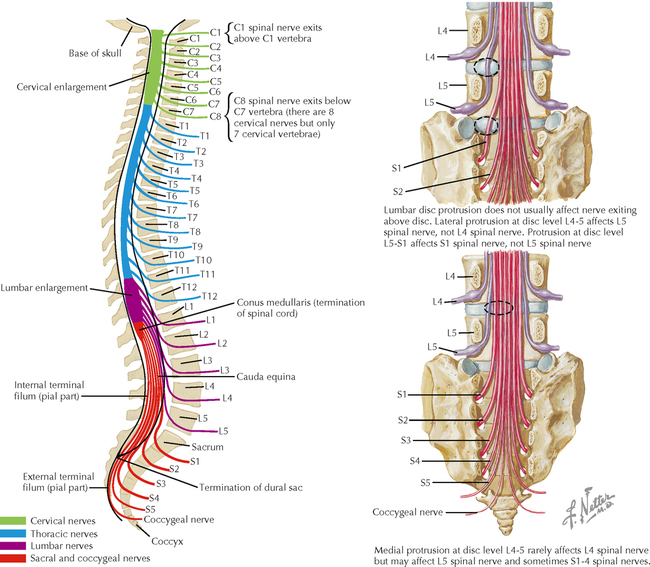
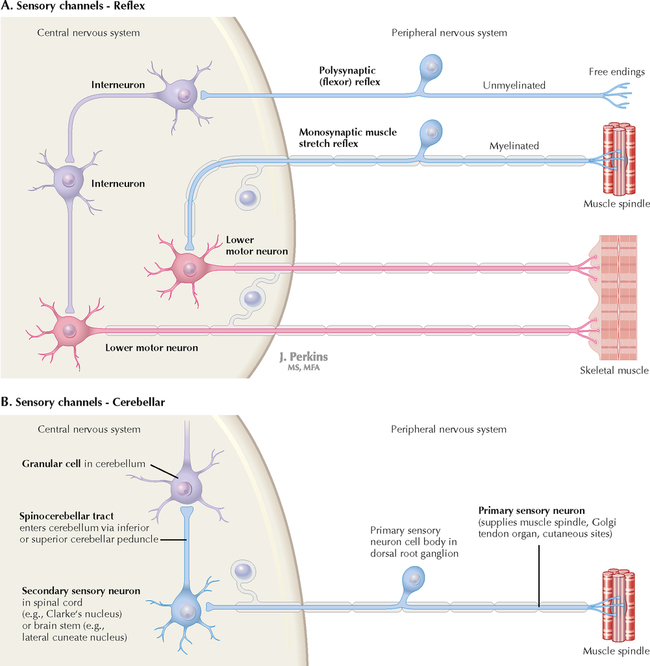
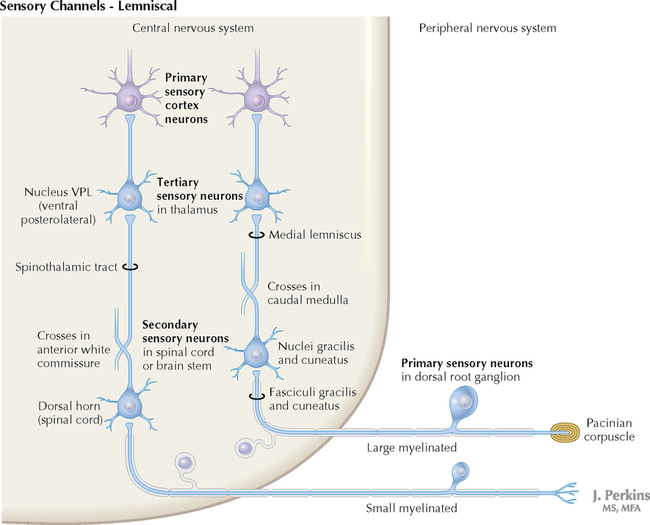
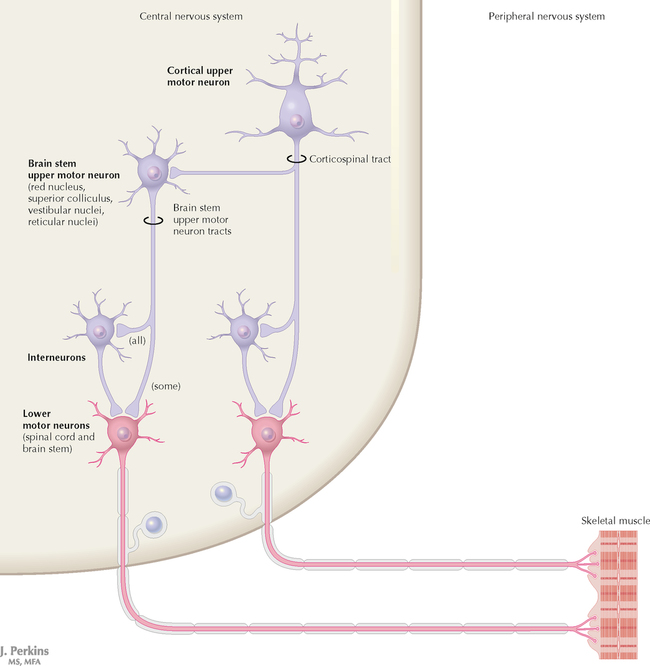
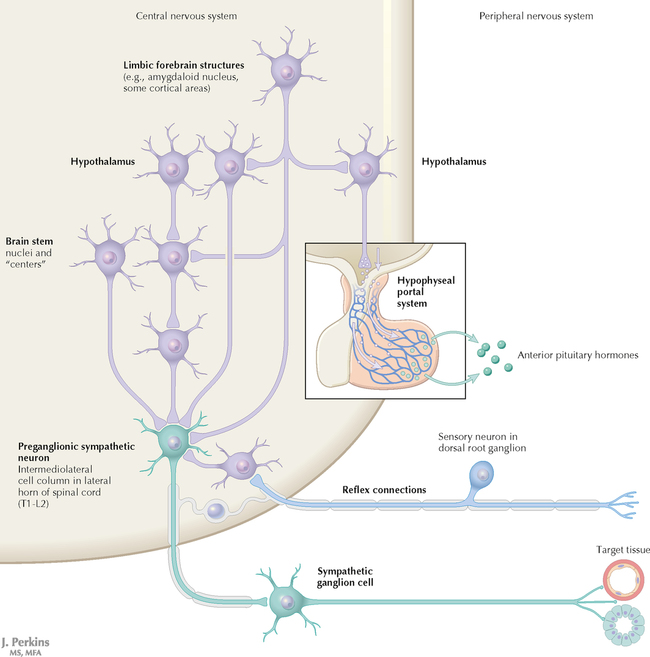
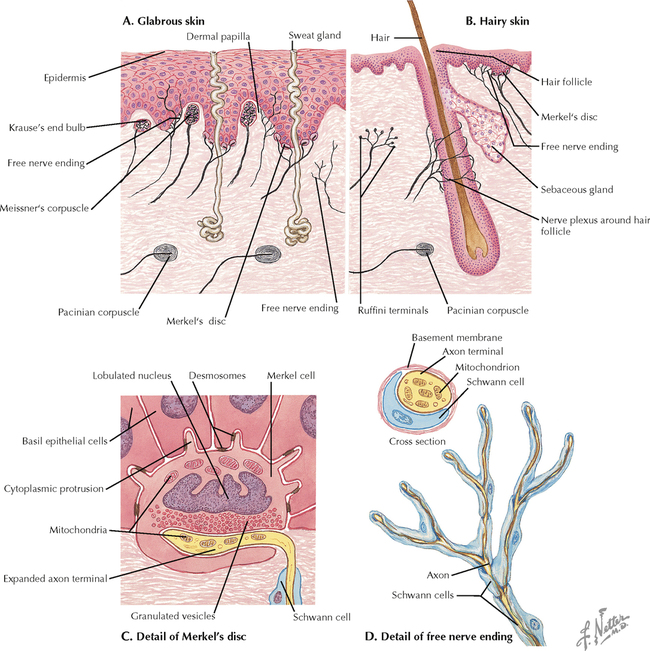
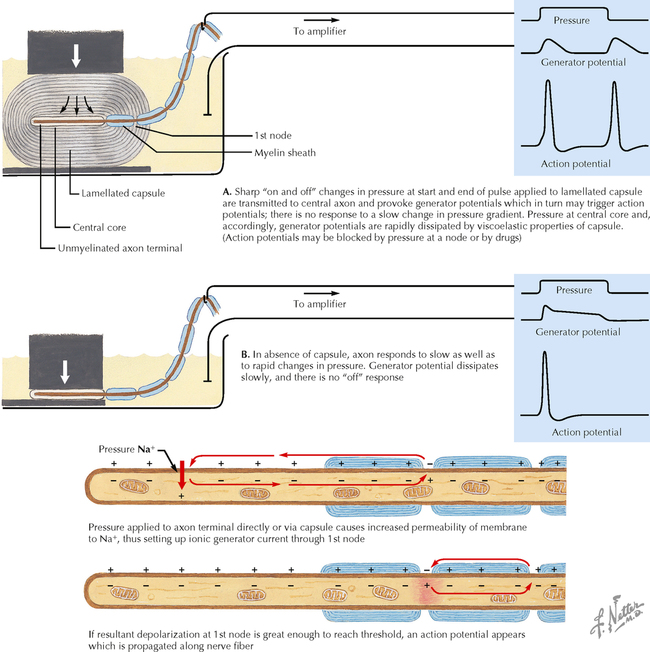
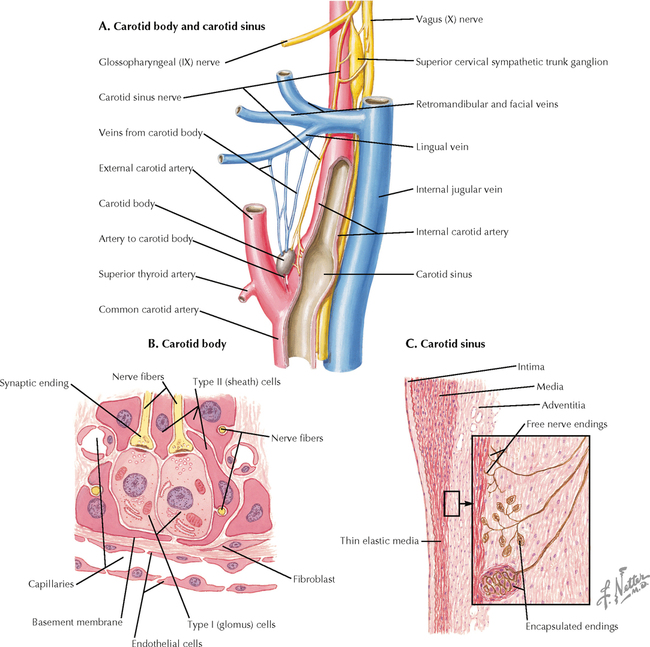
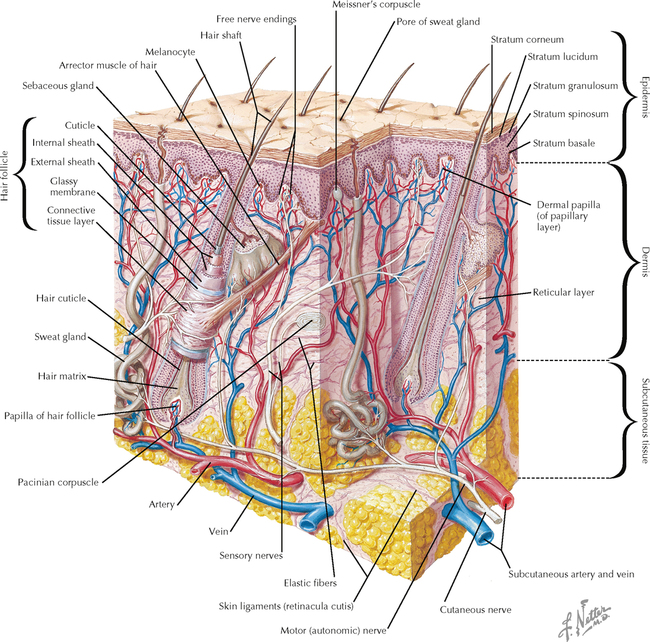
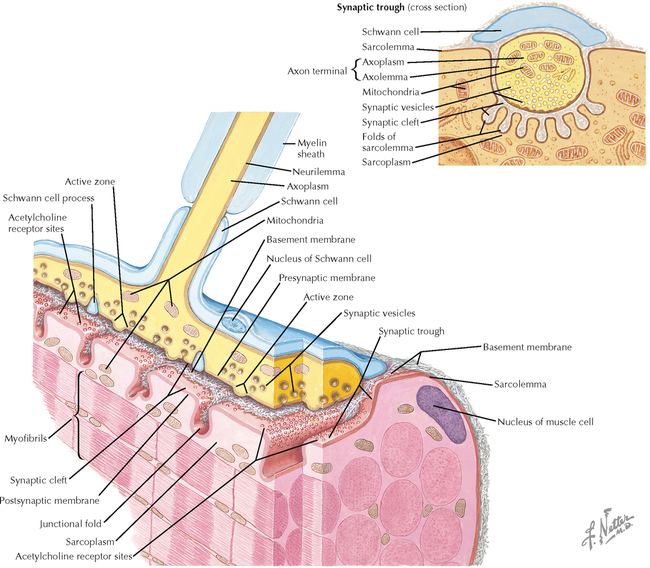
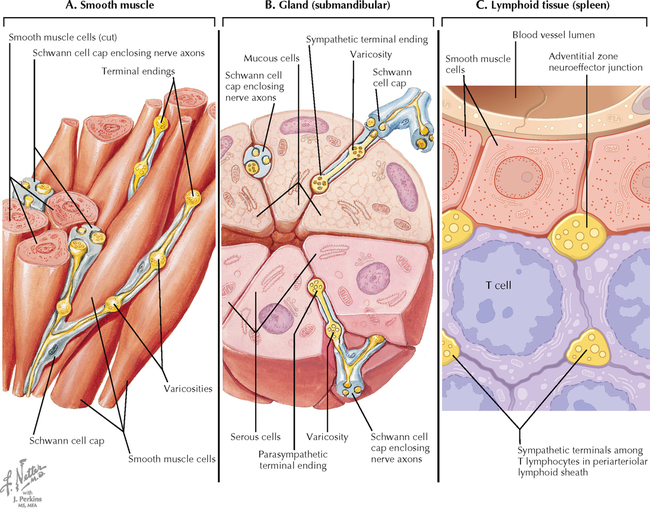
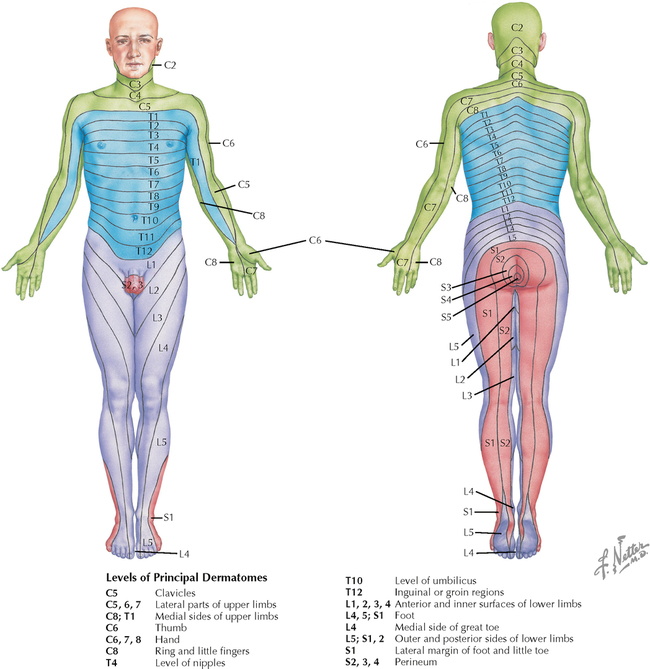
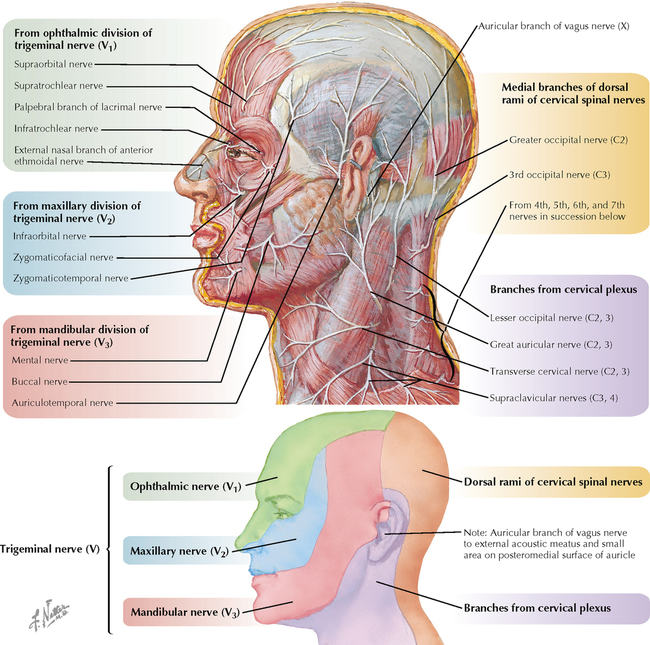
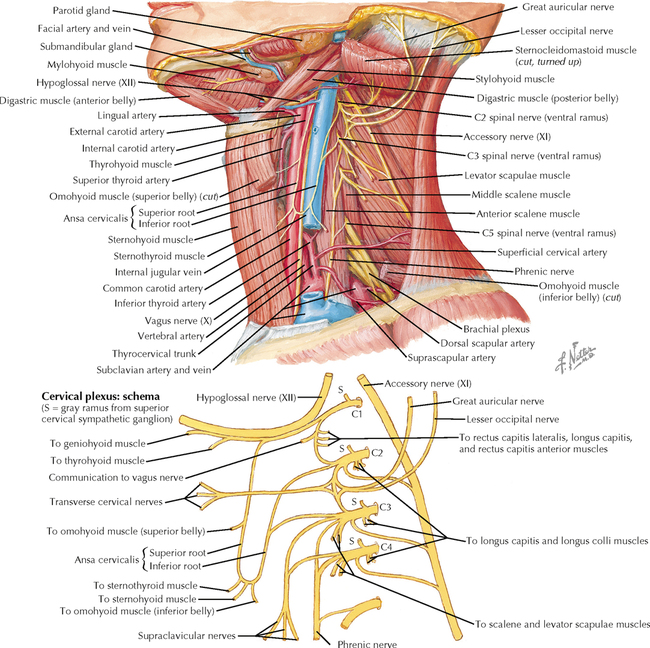
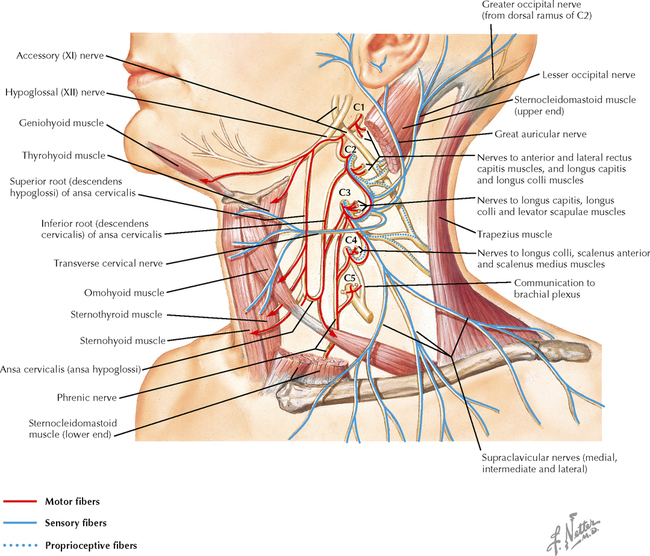
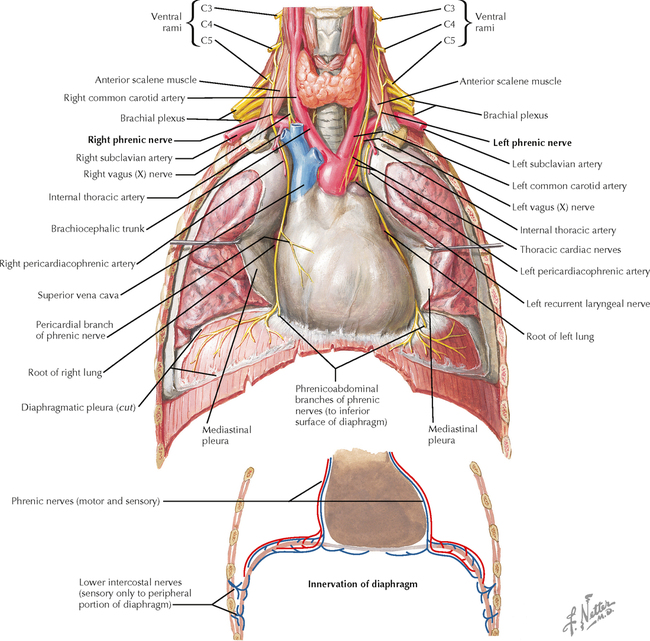
9 PERIPHERAL NERVOUS SYSTEM Introduction and Basic Organization 9.1. Schematic of the Spinal Cord with Sensory, Motor, and Autonomic Components of Peripheral Nerves 9.2. Anatomy of a Peripheral Nerve 9.3. Peripheral Nerve Injury and Degeneration in a Compression Neuropathy 9.4. Relationship of Spinal Nerve Roots to Vertebrae 9.5. Sensory Channels: Reflex and Cerebellar 9.6. Sensory Channels: Lemniscal 9.7. Motor Channels: Basic Organization of Lower and Upper Motor Neurons 9.8. Autonomic Channels 9.9. Cutaneous Receptors 9.10. Pacinian Corpuscles 9.11. Interoceptors 9.12. Skin and Its Nerves 9.13. The Neuromuscular Junction and Neurotransmission 9.14. Neuroeffector Junctions Somatic Nervous System 9.15. Dermatomal Distribution 9.16. Cutaneous Nerves of the Head and Neck 9.17. Cervical Plexus in Situ 9.18. Cervical Plexus 9.19. Phrenic Nerve 9.20. Thoracic Nerves 9.21. Brachial Plexus 9.22. Dermatomes of the Upper Limb 9.23. Cutaneous Innervation of the Upper Limb 9.24. The Scapular, Axillary, and Radial Nerves above the Elbow 9.25. Radial Nerve in the Forearm 9.26. Musculocutaneous Nerve 9.27. Median Nerve 9.28. Carpal Tunnel Syndrome 9.29. Ulnar Nerve 9.30. Lumbar Plexus 9.31. Sacral and Coccygeal Plexuses 9.32. Femoral and Lateral Femoral Cutaneous Nerves 9.33. Obturator Nerve 9.34. Sciatic and Posterior Femoral Cutaneous Nerves 9.35. Tibial Nerve 9.36. Common Peroneal Nerve Autonomic Nervous System 9.37. General Schema of the Autonomic Nervous System 9.38. Autonomic Innervation of Organs of the Immune System and Metabolic Organs 9.39. Reflex Pathways 9.40. Cholinergic and Adrenergic Synapses 9.41. Schematic of Cholinergic and Adrenergic Distribution to Motor and Autonomic Structures 9.42. Autonomic Distribution to the Head and Neck: Medial View 9.43. Autonomic Distribution to the Head and Neck: Lateral View 9.44. Schematic of Autonomic Distribution to the Head and Neck 9.45. Autonomic Distribution to the Eye 9.46. Autonomic Innervation of the Nasal Cavity 9.47. Schematic of the Pterygopalatine and Submandibular Ganglia 9.48. Schematic of the Otic Ganglion 9.49. Innervation of the Limbs 9.50. Thoracic Sympathetic Chain and Splanchnic Nerves 9.51. Innervation of the Tracheobronchial Tree 9.52. Innervation of the Heart 9.53. Abdominal Nerves and Ganglia 9.54. Nerves of the Esophagus 9.55. Innervation of the Stomach and Proximal Duodenum 9.56. Nerves of the Stomach and Duodenum 9.57. Innervation of the Small and Large Intestines 9.58. Nerves of the Small Intestine 9.59. Nerves of the Large Intestine 9.60. Enteric Nervous System: Longitudinal View 9.61. Enteric Nervous System: Cross-sectional View 9.62. Autonomic Innervation of the Liver and Biliary Tract 9.63. Autonomic Innervation of the Pancreas 9.64. Schematic of Innervation of the Adrenal Gland 9.65. Innervation of the Adrenal Gland 9.66. Autonomic Pelvic Nerves and Ganglia 9.67. Nerves of the Kidneys, Ureters, and Urinary Bladder 9.68. Innervation of the Kidney and Upper Ureter 9.69. Innervation of the Urinary Bladder and Lower Ureter 9.70. Innervation of the Male Reproductive Organs 9.71. Innervation of the Female Reproductive Organs INTRODUCTION AND BASIC ORGANIZATION 9.1 SCHEMATIC OF THE SPINAL CORD WITH SENSORY, MOTOR, AND AUTONOMIC COMPONENTS OF PERIPHERAL NERVES Peripheral nerves consist of axons from primary sensory neurons, lower motor neurons (LMNs), and preganglionic and postganglionic autonomic neurons. The primary sensory axons have sensory receptors (transducing elements) at their peripheral (distal) ends, contiguous with the initial segment of the axon. The proximal portion of the axon enters the central nervous system (CNS) and terminates in secondary sensory nuclei associated with reflex, cerebellar, and lemniscal channels. LMNs in the anterior horn of the spinal cord send axons via the ventral (anterior) roots to travel in peripheral nerves to skeletal muscles, with which they form neuromuscular junctions. The autonomic preganglionic neurons send axons via the ventral roots to terminate in autonomic ganglia or in the adrenal medulla. Postganglionic neurons send axons into splanchnic or peripheral nerves and form neuroeffector junctions with smooth muscle, cardiac muscle, secretory glands, metabolic cells, and cells of the immune system. CLINICAL POINT Peripheral nerves form through the union of dorsal and ventral roots and by subsequent branching, similar to the process that occurs through the brachial plexus. The resultant terminal peripheral nerves contain limited categories of axonal types, including LMN axons (both alpha and gamma); primary sensory axons (both myelinated and unmyelinated); and autonomic axons (mainly postganglionic sympathetic axons). Destructive lesions in peripheral nerves may cause flaccid paralysis of innervated skeletal muscles (with loss of tone and denervation atrophy); loss of some or all aspects of somatic sensation in the innervated territory; and some autonomic dysfunction resulting from loss of sympathetic innervation (e.g., vasodilation and lack of sweating). An irritative lesion of a peripheral nerve is usually manifested as pain radiating to the innervated territory. 9.2 ANATOMY OF A PERIPHERAL NERVE A peripheral nerve is made up of unmyelinated and myelinated axons; the connective sheaths with which they are associated; and local blood vessels, the vasa nervorum. Unmyelinated axons are surrounded by the cytoplasm of Schwann cells, called Schwann cell sheaths. Each individual segment of a myelinated axon is enwrapped by a myelin sheath, provided by an individual Schwann cell. The bare space between each myelin sheath is called a node of Ranvier and is the site on the membrane where sodium channels are present and is also the site of initiation or reinitiation of the action potential. Endoneurium is loose, supportive, connective tissue that is found between individual axons within a fascicle. Fascicles of multiple axons are enwrapped by a sheath of supportive cells and collagenous connective tissue; this perineurium functions as a blood-nerve barrier and helps to protect the axons from local diffusion of potentially damaging substances. This perineurial barrier can be disrupted in neuropathic conditions such as diabetic neuropathy. The epineurium is the outermost layer of supportive connective tissue that enwraps the entire nerve. 9.3 PERIPHERAL NERVE INJURY AND DEGENERATION IN A COMPRESSION NEUROPATHY If a peripheral nerve is compressed or damaged, a series of reactions takes place within the neurons whose axons have been damaged and in the supportive tissue. At the site of the injury, axonal damage and thinning of the myelination can occur. Distal to the site of the injury, the peripheral portion of the axon can degenerate (called Wallerian degeneration), resulting in the breaking up and dissolution of the peripheral axon. The Schwann cells myelinating the degenerating axons also break up and degenerate. However, the basement membrane remains intact, providing a scaffold through which future regenerating axons can be directed. The central (proximal) portion of the neuron can undergo changes called central chromatolysis. The Nissl bodies break up into individual ribosomes, the cell body swells, and the neuron shifts its metabolism to structural and reparative synthetic products that attempt to save the neuron and permit it to try to recover from the injury. If successful, this process gradually reverses, and the neuron begins to sprout a peripheral axonal extension, seeking to reattach to the target from which it was disrupted. The Schwann cells proliferate and generate new myelin sheaths around the regrowing axon, but the intersegmental distances of the new myelin sheath are shorter than the original distances and the myelin sheath is thinner; thus, the regenerated axon shows a slower conduction velocity than the original intact axon. 9.4 RELATIONSHIP OF SPINAL NERVE ROOTS TO VERTEBRAE The dorsal (posterior) and ventral (anterior) roots of the spinal cord segments extend from the spinal cord as peripheral axons, invested initially with meninges. As the axons enter the peripheral nervous system, they become associated with Schwann cells for myelination and support. The roots exit through the intervertebral foramina, compact openings between the vertebrae where herniated discs (nucleus pulposus) may impinge on the nerve roots and produce sensory or motor symptoms. Sensory and motor axons enter into the dorsal and ventral rami of peripheral nerves. Autonomic axons (myelinated) course from the ventral roots into the white (preganglionic) rami communicantes and synapse in autonomic ganglia. The ganglion cells give rise to postganglionic axons (unmyelinated) that course through the gray rami communicantes and join the peripheral nerves. CLINICAL POINT The longitudinal growth of the spinal column outstrips the longitudinal growth of the spinal cord; as a consequence, the spinal cord in adults ends adjacent to the L1 vertebral body. Nerve roots heading for intervertebral foramina below L1 extend caudally through the subarachnoid space in the lumbar cistern, forming the cauda equina. Damage to the cauda equina can occur as the result of tumors, such as ependymomas and lipomas, or of a prolapsed intervertebral disc. It is common for symptoms to occur gradually and be irregular because of the ample room in the lumbar cistern for nerve roots to move. Radicular pain often is experienced in a sciatic distribution, with progressive loss of sensation in radicular patterns. A more caudal location of the obstructing mass may lead to loss of sensation in regions of sacral innervation in the perineal (saddle) zone. Loss of bowel, bladder, and erectile function also may occur. More rostral lesions may result in flaccid paralysis of the legs. 9.5 SENSORY CHANNELS: REFLEX AND CEREBELLAR Primary sensory axons communicate with secondary sensory neurons in reflex, cerebellar, and lemniscal channels, carrying transduced information from the periphery into the CNS. A, The reflex channels interconnect primary sensory axons with anterior horn cells (LMNs) through one or more synapses to achieve unconscious reflex motor responses to sensory input. These responses can be elicited in an isolated spinal cord devoid of connections from the brain. The monosynaptic reflex channels connect primary sensory axons from muscle spindles, via the dorsal roots, directly with LMNs involved in muscle stretch reflex contraction; this is the only monosynaptic reflex seen in the human CNS. Polysynaptic reflex channels are directed particularly toward flexor (withdrawal) responses through one or more interneurons to produce coordinated patterns of muscle activity to remove a portion of the body from a potentially damaging or offending stimulus. This polysynaptic channel can spread ipsilaterally and contralaterally through many segments. B, Primary somatosensory axons carrying unconsciously processed information from muscles, joints, tendons, ligaments, and cutaneous sources enter the CNS via dorsal roots and synapse with secondary sensory neurons in the spinal cord or caudal brain stem. These secondary sensory neurons convey information, initially derived from the periphery, to the ipsilateral cerebellum via spinocerebellar pathways. The dorsal and ventral spinocerebellar pathways carry information from the lower body (T6 and below). The rostral spinocerebellar tract and the cuneocerebellar tract carry information from the upper body (above T6). Polysynaptic indirect spinocerebellar pathways (spino-olivo-cerebellar and spinoreticulo-cerebellar tracts) also are present. 9.6 SENSORY CHANNELS: LEMNISCAL Primary sensory axons carrying sensory information destined for conscious perception arise from receptors in superficial and deep tissue. These axons enter the CNS via the dorsal roots and terminate on secondary sensory nuclei in the spinal cord or brain stem. Secondary sensory axons from these nuclei cross the midline (decussate), ascend as lemniscal pathways, and terminate in the contralateral thalamus. These specific thalamic nuclei then project to specific regions of the primary sensory cortex, where fine-grained analysis of incoming, consciously perceived sensory information takes place. Somatosensory information is directed into two sets of channels, protopathic and epicritic. The epicritic information (fine, discriminative sensation; vibratory sensation; joint position sense) is transduced by primary sensory neurons (dorsal root ganglion cells) that send myelinated axons to neurons in the medulla; the nucleus gracilis (lower body, T6 and below); and the nucleus cuneatus (upper body, above T6). Nuclei gracilis and cuneatus give rise to the medial lemniscus, a crossed secondary sensory pathway that terminates in the ventral posterolateral nucleus of the thalamus. This thalamic nucleus has reciprocal projections with cortical neurons in the postcentral gyrus (Brodmann’s areas 3, 1, and 2). This entire epicritic somatosensory system is highly topographically organized, with each region of the body represented in each nucleus and axonal pathway. The protopathic information (pain, temperature sensation, light moving touch) is transduced by primary sensory neurons (dorsal root ganglion cells) that project mainly via small myelinated and unmyelinated axons to neurons in the dorsal horn of the spinal cord. These spinal cord neurons give rise to the spinothalamic tract (spinal lemniscus), a secondary sensory pathway that terminates in separate neuronal sites in the ventral posterolateral nucleus of the thalamus. This portion of the ventral posterolateral nucleus communicates mainly with the primary sensory cortex (SI) and a secondary area of somatosensory cortex (SII) posterior to the lateral postcentral gyrus. Some unmyelinated nociceptive protopathic axons that terminate in the dorsal horn of the spinal cord interconnect with a cascade of spinal cord interneurons that project mainly into the reticular formation of the brain stem (the spinoreticular pathway). This more diffuse pain system is processed through nonspecific thalamic nuclei with projections to somatosensory cortices and more widespread regions of cortex. This system can result in the perception of excruciating, long-lasting pain that may exceed the duration and intensity of direct peripheral stimuli. 9.7 MOTOR CHANNELS: BASIC ORGANIZATION OF LOWER AND UPPER MOTOR NEURONS LMNs are found in the anterior horn of the spinal cord and in motor cranial nerve nuclei in the brain stem. Their axons exit via the ventral roots or cranial nerves to supply skeletal muscles. LMN synapses with muscle fibers form neuromuscular junctions and release the neurotransmitter acetylcholine, which acts on nicotinic receptors on the skeletal muscle fibers. A motor unit consists of an LMN, its axon, and the muscle fibers the axon innervates. LMNs are regulated and coordinated by groups of upper motor neurons (UMNs) found in the brain. Brain stem UMNs regulate basic tone and posture. Cortical UMNs (from corticospinal and corticobulbar tracts) regulate consciously directed, or volitional, movements. Cortical UMNs also have extensive connections with brain stem UMNs and may help to coordinate their activities. The cerebellum and basal ganglia aid in the coordination of movement and in pattern selection, respectively, via connections with UMNs; the cerebellum and basal ganglia do not connect with LMNs directly. 9.8 AUTONOMIC CHANNELS Preganglionic neurons for the sympathetic nervous system (SNS) are found in the lateral horn (intermediolateral cell column) of the thoracolumbar (T1–L2) spinal cord (thoracolumbar system). Preganglionic neurons for the parasympathetic nervous system (PsNS) are found in nuclei of cranial nerves (CNs) III, VII, IX, and X and in the intermediate gray matter of the spinal cord between S2 and S4 (the craniosacral system). Preganglionic axons exit the CNS via cranial nerves or ventral roots and terminate in chain ganglia or collateral ganglia (the SNS) or in intramural ganglia in or near the organ innervated (the PsNS). Postganglionic autonomic axons innervate smooth muscle, cardiac muscle, secretory glands, metabolic cells, and cells of the immune system. The SNS is a fight-or-flight system that responds to emergency demands. The PsNS is a homeostatic, reparative system active in more quiescent activities and in digestive and eliminative functions. Preganglionic responses are coordinated by autonomic UMN equivalents from the brain stem (autonomic centers), the hypothalamus, and the limbic forebrain structures. Inputs that affect visceral functions or elicit emotional responsiveness are conveyed through these central autonomic regulatory systems, which help to coordinate appropriate autonomic responses. These central autonomic regulatory systems coordinate autonomic responses that affect both visceral functions and neuroendocrine outflow from the pituitary gland. 9.9 CUTANEOUS RECEPTORS Cutaneous receptors are found at the distal ends of the primary sensory axon; they act as dendrites, in which threshold stimuli lead to the firing of an action potential at the initial segment of the primary sensory axon. Although specific types of sensory receptors are thought to code for consciously perceived perceptions, there is not an exact correlation. Glabrous skin and hairy skin contain a wide variety of sensory receptors for detecting mechanical, thermal, or nociceptive (consciously perceived as painful) stimuli applied on the body surface. These receptors include bare nerve endings (nociception, thermal sensation) and encapsulated endings. The latter include pacinian corpuscles (rapidly adapting mechanoreceptors for detecting vibration or brief touch); Merkel’s discs (slowly adapting mechanoreceptors for detecting maintained deformation or sustained touch on the skin); Meissner’s corpuscles (rapidly adapting mechanoreceptors for detecting moving touch); Ruffini endings (slowly adapting mechanoreceptors for detecting steady pressure applied to hairy skin); hair follicle receptors (rapidly adapting); and Krause end bulbs (possibly thermoreceptors). The initial segment of the primary sensory axon is immediately adjacent to the sensory receptor. 9.10 PACINIAN CORPUSCLES Pacinian corpuscles are mechanoreceptors that transform mechanical force or displacement into action potentials in large-diameter primary sensory axons. The mechanical stimulus is modified by the viscoelastic properties of the contributing lamellae of the pacinian corpuscle and the associated accessory cells. An action potential results when a generator potential of sufficient magnitude to bring the initial segment of the axon to threshold is elicited. The onset and cessation of mechanical deformation enhance ionic permeability in the axon, optimizing the physiological response of the pacinian corpuscle to vibratory stimuli. 9.11 INTEROCEPTORS Interoceptors, including internal nociceptors, chemoreceptors, and stretch receptors, inform the CNS about the internal state of the body. The carotid body, a specialized chemoreceptor for detecting carbon dioxide (in a hypoxic state) or to a lesser extent low blood pH resulting in increased respiration, is associated with afferent axons of CN IX that project to the caudal nucleus solitarius in the medulla. The carotid sinus, a thin-walled region of the carotid artery, contains encapsulated and bare nerve endings that act as stretch receptors. These stretch receptors respond to increased arterial pressure as baroreceptors, send primary afferents to the caudal nucleus solitarius via CN IX, and elicit reflex bradycardia and decrease in blood pressure. 9.12 SKIN AND ITS NERVES The skin is supplied with a variety of receptor types (see Fig. 9.9) that transduce slowly and rapidly adapting mechanical stimuli and deformation into electrical impulses in primary afferent fibers. The bare nerve endings are associated mainly with nociceptors, peripheral arborizations of unmyelinated axons. Some nociceptors and thermoreceptors are associated with small myelinated axons. These axons collectively contribute somatosensory information to the spinothalamic/spinoreticular lemniscal system for protopathic sensation. The more complex encapsulated receptors contribute somatosensory information to the dorsal column/medial lemniscal system for epicritic sensation and are associated with larger myelinated axons. 9.13 THE NEUROMUSCULAR JUNCTION AND NEUROTRANSMISSION Axons of LMNs that synapse on skeletal muscle form expanded terminals called neuromuscular junctions (motor end plates). The motor axon loses its myelin sheath and expands into an extended terminal that resides in a trough in the muscle fiber and is covered by a layer of Schwann cell cytoplasm. The postsynaptic membrane is thrown into secondary folds. When an action potential invades the motor terminal, several hundred vesicles simultaneously release their acetylcholine (ACh) into the synaptic cleft. The ACh binds to nicotinic receptors on the muscle sarcolemma, initiating a motor end-plate potential, which is normally of sufficient magnitude to result in the firing of a muscle action potential, leading to contraction of the muscle fiber. A single muscle fiber has only one neuromuscular junction, but a motor axon may innervate multiple muscle fibers. CLINICAL POINT An action potential that invades the motor end plate results in a calcium-mediated simultaneous release of multiple quanta (vesicles) of ACh. This released ACh acts on nicotinic cholinergic receptors on the postjunctional membrane, normally resulting in a muscle contraction (excitation-contraction coupling). In myasthenia gravis, antibodies against the cholinergic nicotinic receptors greatly reduce the number of active receptors available for stimulation by released ACh. The size and number of ACh quanta appear to be normal. As a consequence, there is easy fatiguability of involved muscles with repeated attempts at contraction. Ocular, facial, and bulbar muscles are the most likely to be affected by this disease, with resultant ptosis, drooping face, diplopia with strabismus, and dysarthria, dysphonia, and dysphagia. Limb muscles (mainly proximal) are involved only in advanced myasthenia gravis. The muscles do not show wasting and atrophy because they are not denervated; muscle stretch reflexes are elicitable. 9.14 NEUROEFFECTOR JUNCTIONS Autonomic postganglionic axons form neuroeffector junctions with cardiac muscle, smooth muscle (A), secretory glands (B), metabolic cells such as hepatocytes and fat cells, and cells of the immune system (C). These nerve endings use mainly norepinephrine for the SNS and acetylcholine for the PsNS. These endings do not form classic CNS or motor end-plate synapses; instead, they terminate as neuroeffector junctions, releasing neurotransmitter into interstitial spaces. This permits a widespread diffusion of the neurotransmitter as a paracrine secretion, initiating postsynaptic responses on cells with appropriate receptors (including many types of cells of the immune system). Some close appositions also are found, such as SNS endings on lymphocytes. Not all smooth muscle cells are innervated by neuroeffector junctions; they are coupled by gap junctions and can contract together when the innervated smooth muscle cell contracts. SOMATIC NERVOUS SYSTEM 9.15 DERMATOMAL DISTRIBUTION A dermatome is the cutaneous area supplied by a single spinal nerve root; the cell bodies are located in dorsal root ganglia. The spinal nerve roots are distributed to structures according to their associations with spinal cord segments. The nerve roots supplying neighboring dermatomes overlap. Thus, sectioning or dysfunction of a single dorsal root produces hypoesthesia (diminished sensation), not anesthesia (total loss of sensation) in the region supplied predominantly by that dermatome, as shown in the figure. Dermatomal anesthesia requires damage to at least three dorsal roots: the central dorsal root and the roots above and below it. In contrast, an irritative lesion such as a herniated intervertebral disc may produce sharp, radiating pain within the distribution of the affected dermatome. As the limb buds for the lower extremities develop, they draw out the nerve roots that correspond with their mesodermal cores and ectodermal coverings. The developing lower limbs rotate medially around a longitudinal axis, with a resultant oblique orientation of the dermatomes. The L1 and L2 dermatomes can be found in sites adjacent to S2 and S3 dermatomes because of the intervening segments migrating into more distal parts of the lower limbs. Knowledge of dermatomes is important for localizing peripheral nerve root lesions and distinguishing them from peripheral nerve lesions. 9.16 CUTANEOUS NERVES OF THE HEAD AND NECK Cutaneous nerves of the head and neck derive from dorsal rami of cervical spinal nerves, from branches from the cervical plexus, and from all three divisions of the trigeminal nerve (CN V). 9.17 CERVICAL PLEXUS IN SITU This diagram of the cervical plexus in situ and the schema below demonstrate the distribution of branches from the C1–C4 nerve roots into the associated peripheral nerves and branches to the innervated muscles. 9.18 CERVICAL PLEXUS The cervical plexus lies deep to the sternocleidomastoid muscle. Its branches convey motor fibers to many cervical muscles and to the diaphragm. Its sensory fibers convey exteroceptive information from parts of the scalp, neck, and chest as well as proprioceptive information from muscles, tendons, and joints. Sympathetic sudomotor and vasomotor fibers travel with this plexus to blood vessels and glands. The superficial branches perforate the cervical fascia to supply cutaneous structures; the deep branches supply mainly muscles and joints. CLINICAL POINT The cervical plexus is formed from the anterior primary rami of C1–C4, deep to the sternocleidomastoid muscle and in front of the scalenus medius and levator scapulae muscles. Sensory branches include the greater and lesser occipital nerves, great auricular nerve, cutaneous cervical nerves, and supraclavicular nerves. The motor branches include the ansa hypoglossi, branches to scalenus medius and levator scapulae muscles, the phrenic nerve, and branches to the spinal accessory nerve. Lesions of the cervical plexus are uncommon, usually resulting from trauma, mass lesions, or as sequelae to surgery such as carotid endartectomy. Involvement of motor branches results in disruption of muscular function, such as shoulder elevation and head rotation and flexion with spinal accessory nerve damage. Involvement of sensory branches results in loss of cutaneous sensation or in pain and paresthesias in regions of the head or neck supplied by these branches. 9.19 PHRENIC NERVE The left and right phrenic nerves are the motor nerves that supply both sides of the diaphragm from the C3, C4, and C5 ventral roots. The phrenic nerve also contains many sensory nerve fibers that supply the fibrous pericardium, the mediastinal pleura, and central areas of the diaphragmatic pleura. Sympathetic postganglionic nerve fibers also travel with this nerve. Coordinated contraction of the diaphragm relies on central control of firing of LMNs through dendrite bundles in the spinal cord. CLINICAL POINT Only gold members can continue reading. Log In or Register to continue Share this:Click to share on Twitter (Opens in new window)Click to share on Facebook (Opens in new window) Related Related posts: SPINAL CORD VENTRICLES AND THE CEREBROSPINAL FLUID SPINAL CORD BRAIN STEM AND CEREBELLUM VASCULATURE DEVELOPMENTAL NEUROSCIENCE Stay updated, free articles. Join our Telegram channel Join Tags: Netters Atlas of Neuroscience with STUDENT CONSULT Online Access Jun 4, 2016 | Posted by admin in NEUROLOGY | Comments Off on PERIPHERAL NERVOUS SYSTEM Full access? Get Clinical Tree
9 PERIPHERAL NERVOUS SYSTEM Introduction and Basic Organization 9.1. Schematic of the Spinal Cord with Sensory, Motor, and Autonomic Components of Peripheral Nerves 9.2. Anatomy of a Peripheral Nerve 9.3. Peripheral Nerve Injury and Degeneration in a Compression Neuropathy 9.4. Relationship of Spinal Nerve Roots to Vertebrae 9.5. Sensory Channels: Reflex and Cerebellar 9.6. Sensory Channels: Lemniscal 9.7. Motor Channels: Basic Organization of Lower and Upper Motor Neurons 9.8. Autonomic Channels 9.9. Cutaneous Receptors 9.10. Pacinian Corpuscles 9.11. Interoceptors 9.12. Skin and Its Nerves 9.13. The Neuromuscular Junction and Neurotransmission 9.14. Neuroeffector Junctions Somatic Nervous System 9.15. Dermatomal Distribution 9.16. Cutaneous Nerves of the Head and Neck 9.17. Cervical Plexus in Situ 9.18. Cervical Plexus 9.19. Phrenic Nerve 9.20. Thoracic Nerves 9.21. Brachial Plexus 9.22. Dermatomes of the Upper Limb 9.23. Cutaneous Innervation of the Upper Limb 9.24. The Scapular, Axillary, and Radial Nerves above the Elbow 9.25. Radial Nerve in the Forearm 9.26. Musculocutaneous Nerve 9.27. Median Nerve 9.28. Carpal Tunnel Syndrome 9.29. Ulnar Nerve 9.30. Lumbar Plexus 9.31. Sacral and Coccygeal Plexuses 9.32. Femoral and Lateral Femoral Cutaneous Nerves 9.33. Obturator Nerve 9.34. Sciatic and Posterior Femoral Cutaneous Nerves 9.35. Tibial Nerve 9.36. Common Peroneal Nerve Autonomic Nervous System 9.37. General Schema of the Autonomic Nervous System 9.38. Autonomic Innervation of Organs of the Immune System and Metabolic Organs 9.39. Reflex Pathways 9.40. Cholinergic and Adrenergic Synapses 9.41. Schematic of Cholinergic and Adrenergic Distribution to Motor and Autonomic Structures 9.42. Autonomic Distribution to the Head and Neck: Medial View 9.43. Autonomic Distribution to the Head and Neck: Lateral View 9.44. Schematic of Autonomic Distribution to the Head and Neck 9.45. Autonomic Distribution to the Eye 9.46. Autonomic Innervation of the Nasal Cavity 9.47. Schematic of the Pterygopalatine and Submandibular Ganglia 9.48. Schematic of the Otic Ganglion 9.49. Innervation of the Limbs 9.50. Thoracic Sympathetic Chain and Splanchnic Nerves 9.51. Innervation of the Tracheobronchial Tree 9.52. Innervation of the Heart 9.53. Abdominal Nerves and Ganglia 9.54. Nerves of the Esophagus 9.55. Innervation of the Stomach and Proximal Duodenum 9.56. Nerves of the Stomach and Duodenum 9.57. Innervation of the Small and Large Intestines 9.58. Nerves of the Small Intestine 9.59. Nerves of the Large Intestine 9.60. Enteric Nervous System: Longitudinal View 9.61. Enteric Nervous System: Cross-sectional View 9.62. Autonomic Innervation of the Liver and Biliary Tract 9.63. Autonomic Innervation of the Pancreas 9.64. Schematic of Innervation of the Adrenal Gland 9.65. Innervation of the Adrenal Gland 9.66. Autonomic Pelvic Nerves and Ganglia 9.67. Nerves of the Kidneys, Ureters, and Urinary Bladder 9.68. Innervation of the Kidney and Upper Ureter 9.69. Innervation of the Urinary Bladder and Lower Ureter 9.70. Innervation of the Male Reproductive Organs 9.71. Innervation of the Female Reproductive Organs INTRODUCTION AND BASIC ORGANIZATION 9.1 SCHEMATIC OF THE SPINAL CORD WITH SENSORY, MOTOR, AND AUTONOMIC COMPONENTS OF PERIPHERAL NERVES Peripheral nerves consist of axons from primary sensory neurons, lower motor neurons (LMNs), and preganglionic and postganglionic autonomic neurons. The primary sensory axons have sensory receptors (transducing elements) at their peripheral (distal) ends, contiguous with the initial segment of the axon. The proximal portion of the axon enters the central nervous system (CNS) and terminates in secondary sensory nuclei associated with reflex, cerebellar, and lemniscal channels. LMNs in the anterior horn of the spinal cord send axons via the ventral (anterior) roots to travel in peripheral nerves to skeletal muscles, with which they form neuromuscular junctions. The autonomic preganglionic neurons send axons via the ventral roots to terminate in autonomic ganglia or in the adrenal medulla. Postganglionic neurons send axons into splanchnic or peripheral nerves and form neuroeffector junctions with smooth muscle, cardiac muscle, secretory glands, metabolic cells, and cells of the immune system. CLINICAL POINT Peripheral nerves form through the union of dorsal and ventral roots and by subsequent branching, similar to the process that occurs through the brachial plexus. The resultant terminal peripheral nerves contain limited categories of axonal types, including LMN axons (both alpha and gamma); primary sensory axons (both myelinated and unmyelinated); and autonomic axons (mainly postganglionic sympathetic axons). Destructive lesions in peripheral nerves may cause flaccid paralysis of innervated skeletal muscles (with loss of tone and denervation atrophy); loss of some or all aspects of somatic sensation in the innervated territory; and some autonomic dysfunction resulting from loss of sympathetic innervation (e.g., vasodilation and lack of sweating). An irritative lesion of a peripheral nerve is usually manifested as pain radiating to the innervated territory. 9.2 ANATOMY OF A PERIPHERAL NERVE A peripheral nerve is made up of unmyelinated and myelinated axons; the connective sheaths with which they are associated; and local blood vessels, the vasa nervorum. Unmyelinated axons are surrounded by the cytoplasm of Schwann cells, called Schwann cell sheaths. Each individual segment of a myelinated axon is enwrapped by a myelin sheath, provided by an individual Schwann cell. The bare space between each myelin sheath is called a node of Ranvier and is the site on the membrane where sodium channels are present and is also the site of initiation or reinitiation of the action potential. Endoneurium is loose, supportive, connective tissue that is found between individual axons within a fascicle. Fascicles of multiple axons are enwrapped by a sheath of supportive cells and collagenous connective tissue; this perineurium functions as a blood-nerve barrier and helps to protect the axons from local diffusion of potentially damaging substances. This perineurial barrier can be disrupted in neuropathic conditions such as diabetic neuropathy. The epineurium is the outermost layer of supportive connective tissue that enwraps the entire nerve. 9.3 PERIPHERAL NERVE INJURY AND DEGENERATION IN A COMPRESSION NEUROPATHY If a peripheral nerve is compressed or damaged, a series of reactions takes place within the neurons whose axons have been damaged and in the supportive tissue. At the site of the injury, axonal damage and thinning of the myelination can occur. Distal to the site of the injury, the peripheral portion of the axon can degenerate (called Wallerian degeneration), resulting in the breaking up and dissolution of the peripheral axon. The Schwann cells myelinating the degenerating axons also break up and degenerate. However, the basement membrane remains intact, providing a scaffold through which future regenerating axons can be directed. The central (proximal) portion of the neuron can undergo changes called central chromatolysis. The Nissl bodies break up into individual ribosomes, the cell body swells, and the neuron shifts its metabolism to structural and reparative synthetic products that attempt to save the neuron and permit it to try to recover from the injury. If successful, this process gradually reverses, and the neuron begins to sprout a peripheral axonal extension, seeking to reattach to the target from which it was disrupted. The Schwann cells proliferate and generate new myelin sheaths around the regrowing axon, but the intersegmental distances of the new myelin sheath are shorter than the original distances and the myelin sheath is thinner; thus, the regenerated axon shows a slower conduction velocity than the original intact axon. 9.4 RELATIONSHIP OF SPINAL NERVE ROOTS TO VERTEBRAE The dorsal (posterior) and ventral (anterior) roots of the spinal cord segments extend from the spinal cord as peripheral axons, invested initially with meninges. As the axons enter the peripheral nervous system, they become associated with Schwann cells for myelination and support. The roots exit through the intervertebral foramina, compact openings between the vertebrae where herniated discs (nucleus pulposus) may impinge on the nerve roots and produce sensory or motor symptoms. Sensory and motor axons enter into the dorsal and ventral rami of peripheral nerves. Autonomic axons (myelinated) course from the ventral roots into the white (preganglionic) rami communicantes and synapse in autonomic ganglia. The ganglion cells give rise to postganglionic axons (unmyelinated) that course through the gray rami communicantes and join the peripheral nerves. CLINICAL POINT The longitudinal growth of the spinal column outstrips the longitudinal growth of the spinal cord; as a consequence, the spinal cord in adults ends adjacent to the L1 vertebral body. Nerve roots heading for intervertebral foramina below L1 extend caudally through the subarachnoid space in the lumbar cistern, forming the cauda equina. Damage to the cauda equina can occur as the result of tumors, such as ependymomas and lipomas, or of a prolapsed intervertebral disc. It is common for symptoms to occur gradually and be irregular because of the ample room in the lumbar cistern for nerve roots to move. Radicular pain often is experienced in a sciatic distribution, with progressive loss of sensation in radicular patterns. A more caudal location of the obstructing mass may lead to loss of sensation in regions of sacral innervation in the perineal (saddle) zone. Loss of bowel, bladder, and erectile function also may occur. More rostral lesions may result in flaccid paralysis of the legs. 9.5 SENSORY CHANNELS: REFLEX AND CEREBELLAR Primary sensory axons communicate with secondary sensory neurons in reflex, cerebellar, and lemniscal channels, carrying transduced information from the periphery into the CNS. A, The reflex channels interconnect primary sensory axons with anterior horn cells (LMNs) through one or more synapses to achieve unconscious reflex motor responses to sensory input. These responses can be elicited in an isolated spinal cord devoid of connections from the brain. The monosynaptic reflex channels connect primary sensory axons from muscle spindles, via the dorsal roots, directly with LMNs involved in muscle stretch reflex contraction; this is the only monosynaptic reflex seen in the human CNS. Polysynaptic reflex channels are directed particularly toward flexor (withdrawal) responses through one or more interneurons to produce coordinated patterns of muscle activity to remove a portion of the body from a potentially damaging or offending stimulus. This polysynaptic channel can spread ipsilaterally and contralaterally through many segments. B, Primary somatosensory axons carrying unconsciously processed information from muscles, joints, tendons, ligaments, and cutaneous sources enter the CNS via dorsal roots and synapse with secondary sensory neurons in the spinal cord or caudal brain stem. These secondary sensory neurons convey information, initially derived from the periphery, to the ipsilateral cerebellum via spinocerebellar pathways. The dorsal and ventral spinocerebellar pathways carry information from the lower body (T6 and below). The rostral spinocerebellar tract and the cuneocerebellar tract carry information from the upper body (above T6). Polysynaptic indirect spinocerebellar pathways (spino-olivo-cerebellar and spinoreticulo-cerebellar tracts) also are present. 9.6 SENSORY CHANNELS: LEMNISCAL Primary sensory axons carrying sensory information destined for conscious perception arise from receptors in superficial and deep tissue. These axons enter the CNS via the dorsal roots and terminate on secondary sensory nuclei in the spinal cord or brain stem. Secondary sensory axons from these nuclei cross the midline (decussate), ascend as lemniscal pathways, and terminate in the contralateral thalamus. These specific thalamic nuclei then project to specific regions of the primary sensory cortex, where fine-grained analysis of incoming, consciously perceived sensory information takes place. Somatosensory information is directed into two sets of channels, protopathic and epicritic. The epicritic information (fine, discriminative sensation; vibratory sensation; joint position sense) is transduced by primary sensory neurons (dorsal root ganglion cells) that send myelinated axons to neurons in the medulla; the nucleus gracilis (lower body, T6 and below); and the nucleus cuneatus (upper body, above T6). Nuclei gracilis and cuneatus give rise to the medial lemniscus, a crossed secondary sensory pathway that terminates in the ventral posterolateral nucleus of the thalamus. This thalamic nucleus has reciprocal projections with cortical neurons in the postcentral gyrus (Brodmann’s areas 3, 1, and 2). This entire epicritic somatosensory system is highly topographically organized, with each region of the body represented in each nucleus and axonal pathway. The protopathic information (pain, temperature sensation, light moving touch) is transduced by primary sensory neurons (dorsal root ganglion cells) that project mainly via small myelinated and unmyelinated axons to neurons in the dorsal horn of the spinal cord. These spinal cord neurons give rise to the spinothalamic tract (spinal lemniscus), a secondary sensory pathway that terminates in separate neuronal sites in the ventral posterolateral nucleus of the thalamus. This portion of the ventral posterolateral nucleus communicates mainly with the primary sensory cortex (SI) and a secondary area of somatosensory cortex (SII) posterior to the lateral postcentral gyrus. Some unmyelinated nociceptive protopathic axons that terminate in the dorsal horn of the spinal cord interconnect with a cascade of spinal cord interneurons that project mainly into the reticular formation of the brain stem (the spinoreticular pathway). This more diffuse pain system is processed through nonspecific thalamic nuclei with projections to somatosensory cortices and more widespread regions of cortex. This system can result in the perception of excruciating, long-lasting pain that may exceed the duration and intensity of direct peripheral stimuli. 9.7 MOTOR CHANNELS: BASIC ORGANIZATION OF LOWER AND UPPER MOTOR NEURONS LMNs are found in the anterior horn of the spinal cord and in motor cranial nerve nuclei in the brain stem. Their axons exit via the ventral roots or cranial nerves to supply skeletal muscles. LMN synapses with muscle fibers form neuromuscular junctions and release the neurotransmitter acetylcholine, which acts on nicotinic receptors on the skeletal muscle fibers. A motor unit consists of an LMN, its axon, and the muscle fibers the axon innervates. LMNs are regulated and coordinated by groups of upper motor neurons (UMNs) found in the brain. Brain stem UMNs regulate basic tone and posture. Cortical UMNs (from corticospinal and corticobulbar tracts) regulate consciously directed, or volitional, movements. Cortical UMNs also have extensive connections with brain stem UMNs and may help to coordinate their activities. The cerebellum and basal ganglia aid in the coordination of movement and in pattern selection, respectively, via connections with UMNs; the cerebellum and basal ganglia do not connect with LMNs directly. 9.8 AUTONOMIC CHANNELS Preganglionic neurons for the sympathetic nervous system (SNS) are found in the lateral horn (intermediolateral cell column) of the thoracolumbar (T1–L2) spinal cord (thoracolumbar system). Preganglionic neurons for the parasympathetic nervous system (PsNS) are found in nuclei of cranial nerves (CNs) III, VII, IX, and X and in the intermediate gray matter of the spinal cord between S2 and S4 (the craniosacral system). Preganglionic axons exit the CNS via cranial nerves or ventral roots and terminate in chain ganglia or collateral ganglia (the SNS) or in intramural ganglia in or near the organ innervated (the PsNS). Postganglionic autonomic axons innervate smooth muscle, cardiac muscle, secretory glands, metabolic cells, and cells of the immune system. The SNS is a fight-or-flight system that responds to emergency demands. The PsNS is a homeostatic, reparative system active in more quiescent activities and in digestive and eliminative functions. Preganglionic responses are coordinated by autonomic UMN equivalents from the brain stem (autonomic centers), the hypothalamus, and the limbic forebrain structures. Inputs that affect visceral functions or elicit emotional responsiveness are conveyed through these central autonomic regulatory systems, which help to coordinate appropriate autonomic responses. These central autonomic regulatory systems coordinate autonomic responses that affect both visceral functions and neuroendocrine outflow from the pituitary gland. 9.9 CUTANEOUS RECEPTORS Cutaneous receptors are found at the distal ends of the primary sensory axon; they act as dendrites, in which threshold stimuli lead to the firing of an action potential at the initial segment of the primary sensory axon. Although specific types of sensory receptors are thought to code for consciously perceived perceptions, there is not an exact correlation. Glabrous skin and hairy skin contain a wide variety of sensory receptors for detecting mechanical, thermal, or nociceptive (consciously perceived as painful) stimuli applied on the body surface. These receptors include bare nerve endings (nociception, thermal sensation) and encapsulated endings. The latter include pacinian corpuscles (rapidly adapting mechanoreceptors for detecting vibration or brief touch); Merkel’s discs (slowly adapting mechanoreceptors for detecting maintained deformation or sustained touch on the skin); Meissner’s corpuscles (rapidly adapting mechanoreceptors for detecting moving touch); Ruffini endings (slowly adapting mechanoreceptors for detecting steady pressure applied to hairy skin); hair follicle receptors (rapidly adapting); and Krause end bulbs (possibly thermoreceptors). The initial segment of the primary sensory axon is immediately adjacent to the sensory receptor. 9.10 PACINIAN CORPUSCLES Pacinian corpuscles are mechanoreceptors that transform mechanical force or displacement into action potentials in large-diameter primary sensory axons. The mechanical stimulus is modified by the viscoelastic properties of the contributing lamellae of the pacinian corpuscle and the associated accessory cells. An action potential results when a generator potential of sufficient magnitude to bring the initial segment of the axon to threshold is elicited. The onset and cessation of mechanical deformation enhance ionic permeability in the axon, optimizing the physiological response of the pacinian corpuscle to vibratory stimuli. 9.11 INTEROCEPTORS Interoceptors, including internal nociceptors, chemoreceptors, and stretch receptors, inform the CNS about the internal state of the body. The carotid body, a specialized chemoreceptor for detecting carbon dioxide (in a hypoxic state) or to a lesser extent low blood pH resulting in increased respiration, is associated with afferent axons of CN IX that project to the caudal nucleus solitarius in the medulla. The carotid sinus, a thin-walled region of the carotid artery, contains encapsulated and bare nerve endings that act as stretch receptors. These stretch receptors respond to increased arterial pressure as baroreceptors, send primary afferents to the caudal nucleus solitarius via CN IX, and elicit reflex bradycardia and decrease in blood pressure. 9.12 SKIN AND ITS NERVES The skin is supplied with a variety of receptor types (see Fig. 9.9) that transduce slowly and rapidly adapting mechanical stimuli and deformation into electrical impulses in primary afferent fibers. The bare nerve endings are associated mainly with nociceptors, peripheral arborizations of unmyelinated axons. Some nociceptors and thermoreceptors are associated with small myelinated axons. These axons collectively contribute somatosensory information to the spinothalamic/spinoreticular lemniscal system for protopathic sensation. The more complex encapsulated receptors contribute somatosensory information to the dorsal column/medial lemniscal system for epicritic sensation and are associated with larger myelinated axons. 9.13 THE NEUROMUSCULAR JUNCTION AND NEUROTRANSMISSION Axons of LMNs that synapse on skeletal muscle form expanded terminals called neuromuscular junctions (motor end plates). The motor axon loses its myelin sheath and expands into an extended terminal that resides in a trough in the muscle fiber and is covered by a layer of Schwann cell cytoplasm. The postsynaptic membrane is thrown into secondary folds. When an action potential invades the motor terminal, several hundred vesicles simultaneously release their acetylcholine (ACh) into the synaptic cleft. The ACh binds to nicotinic receptors on the muscle sarcolemma, initiating a motor end-plate potential, which is normally of sufficient magnitude to result in the firing of a muscle action potential, leading to contraction of the muscle fiber. A single muscle fiber has only one neuromuscular junction, but a motor axon may innervate multiple muscle fibers. CLINICAL POINT An action potential that invades the motor end plate results in a calcium-mediated simultaneous release of multiple quanta (vesicles) of ACh. This released ACh acts on nicotinic cholinergic receptors on the postjunctional membrane, normally resulting in a muscle contraction (excitation-contraction coupling). In myasthenia gravis, antibodies against the cholinergic nicotinic receptors greatly reduce the number of active receptors available for stimulation by released ACh. The size and number of ACh quanta appear to be normal. As a consequence, there is easy fatiguability of involved muscles with repeated attempts at contraction. Ocular, facial, and bulbar muscles are the most likely to be affected by this disease, with resultant ptosis, drooping face, diplopia with strabismus, and dysarthria, dysphonia, and dysphagia. Limb muscles (mainly proximal) are involved only in advanced myasthenia gravis. The muscles do not show wasting and atrophy because they are not denervated; muscle stretch reflexes are elicitable. 9.14 NEUROEFFECTOR JUNCTIONS Autonomic postganglionic axons form neuroeffector junctions with cardiac muscle, smooth muscle (A), secretory glands (B), metabolic cells such as hepatocytes and fat cells, and cells of the immune system (C). These nerve endings use mainly norepinephrine for the SNS and acetylcholine for the PsNS. These endings do not form classic CNS or motor end-plate synapses; instead, they terminate as neuroeffector junctions, releasing neurotransmitter into interstitial spaces. This permits a widespread diffusion of the neurotransmitter as a paracrine secretion, initiating postsynaptic responses on cells with appropriate receptors (including many types of cells of the immune system). Some close appositions also are found, such as SNS endings on lymphocytes. Not all smooth muscle cells are innervated by neuroeffector junctions; they are coupled by gap junctions and can contract together when the innervated smooth muscle cell contracts. SOMATIC NERVOUS SYSTEM 9.15 DERMATOMAL DISTRIBUTION A dermatome is the cutaneous area supplied by a single spinal nerve root; the cell bodies are located in dorsal root ganglia. The spinal nerve roots are distributed to structures according to their associations with spinal cord segments. The nerve roots supplying neighboring dermatomes overlap. Thus, sectioning or dysfunction of a single dorsal root produces hypoesthesia (diminished sensation), not anesthesia (total loss of sensation) in the region supplied predominantly by that dermatome, as shown in the figure. Dermatomal anesthesia requires damage to at least three dorsal roots: the central dorsal root and the roots above and below it. In contrast, an irritative lesion such as a herniated intervertebral disc may produce sharp, radiating pain within the distribution of the affected dermatome. As the limb buds for the lower extremities develop, they draw out the nerve roots that correspond with their mesodermal cores and ectodermal coverings. The developing lower limbs rotate medially around a longitudinal axis, with a resultant oblique orientation of the dermatomes. The L1 and L2 dermatomes can be found in sites adjacent to S2 and S3 dermatomes because of the intervening segments migrating into more distal parts of the lower limbs. Knowledge of dermatomes is important for localizing peripheral nerve root lesions and distinguishing them from peripheral nerve lesions. 9.16 CUTANEOUS NERVES OF THE HEAD AND NECK Cutaneous nerves of the head and neck derive from dorsal rami of cervical spinal nerves, from branches from the cervical plexus, and from all three divisions of the trigeminal nerve (CN V). 9.17 CERVICAL PLEXUS IN SITU This diagram of the cervical plexus in situ and the schema below demonstrate the distribution of branches from the C1–C4 nerve roots into the associated peripheral nerves and branches to the innervated muscles. 9.18 CERVICAL PLEXUS The cervical plexus lies deep to the sternocleidomastoid muscle. Its branches convey motor fibers to many cervical muscles and to the diaphragm. Its sensory fibers convey exteroceptive information from parts of the scalp, neck, and chest as well as proprioceptive information from muscles, tendons, and joints. Sympathetic sudomotor and vasomotor fibers travel with this plexus to blood vessels and glands. The superficial branches perforate the cervical fascia to supply cutaneous structures; the deep branches supply mainly muscles and joints. CLINICAL POINT The cervical plexus is formed from the anterior primary rami of C1–C4, deep to the sternocleidomastoid muscle and in front of the scalenus medius and levator scapulae muscles. Sensory branches include the greater and lesser occipital nerves, great auricular nerve, cutaneous cervical nerves, and supraclavicular nerves. The motor branches include the ansa hypoglossi, branches to scalenus medius and levator scapulae muscles, the phrenic nerve, and branches to the spinal accessory nerve. Lesions of the cervical plexus are uncommon, usually resulting from trauma, mass lesions, or as sequelae to surgery such as carotid endartectomy. Involvement of motor branches results in disruption of muscular function, such as shoulder elevation and head rotation and flexion with spinal accessory nerve damage. Involvement of sensory branches results in loss of cutaneous sensation or in pain and paresthesias in regions of the head or neck supplied by these branches. 9.19 PHRENIC NERVE The left and right phrenic nerves are the motor nerves that supply both sides of the diaphragm from the C3, C4, and C5 ventral roots. The phrenic nerve also contains many sensory nerve fibers that supply the fibrous pericardium, the mediastinal pleura, and central areas of the diaphragmatic pleura. Sympathetic postganglionic nerve fibers also travel with this nerve. Coordinated contraction of the diaphragm relies on central control of firing of LMNs through dendrite bundles in the spinal cord. CLINICAL POINT Only gold members can continue reading. Log In or Register to continue Share this:Click to share on Twitter (Opens in new window)Click to share on Facebook (Opens in new window) Related Related posts: SPINAL CORD VENTRICLES AND THE CEREBROSPINAL FLUID SPINAL CORD BRAIN STEM AND CEREBELLUM VASCULATURE DEVELOPMENTAL NEUROSCIENCE Stay updated, free articles. Join our Telegram channel Join Tags: Netters Atlas of Neuroscience with STUDENT CONSULT Online Access Jun 4, 2016 | Posted by admin in NEUROLOGY | Comments Off on PERIPHERAL NERVOUS SYSTEM Full access? Get Clinical Tree