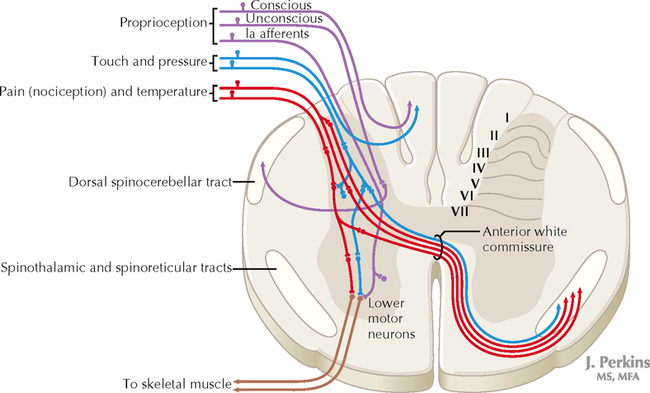
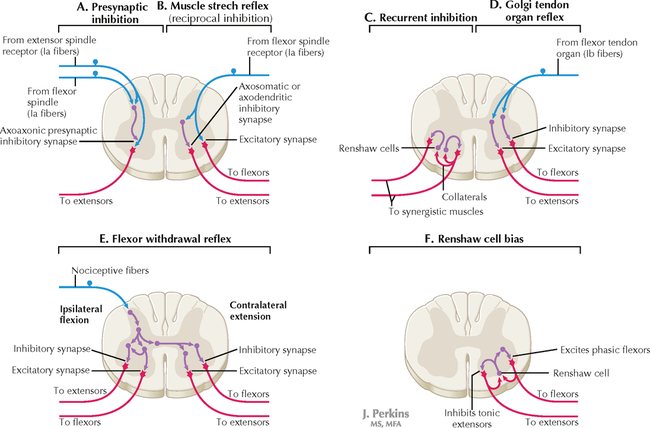
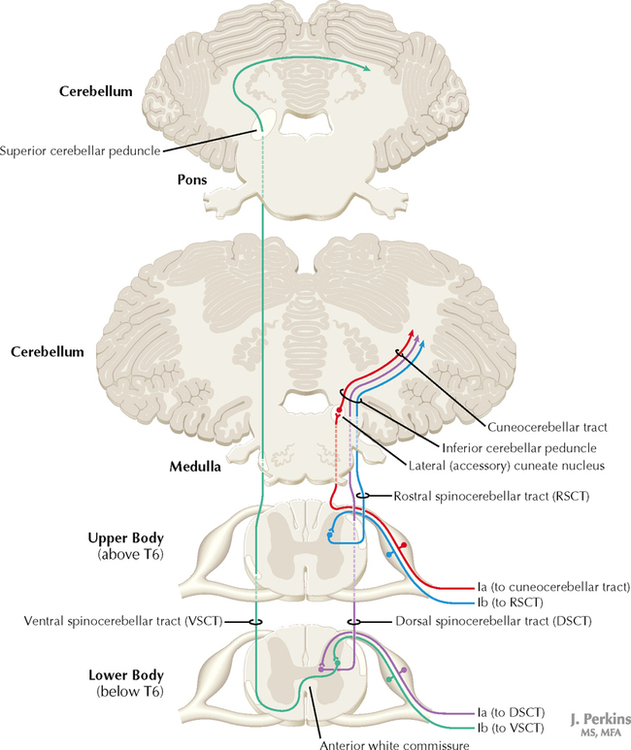
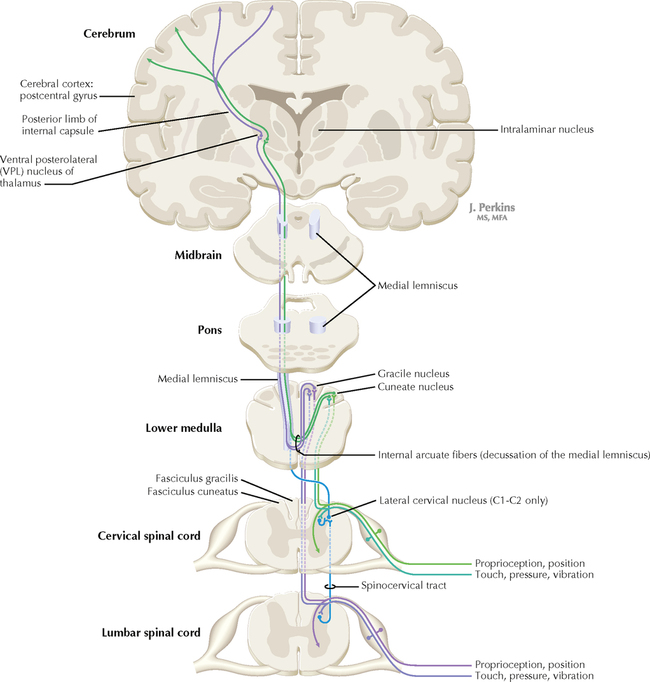
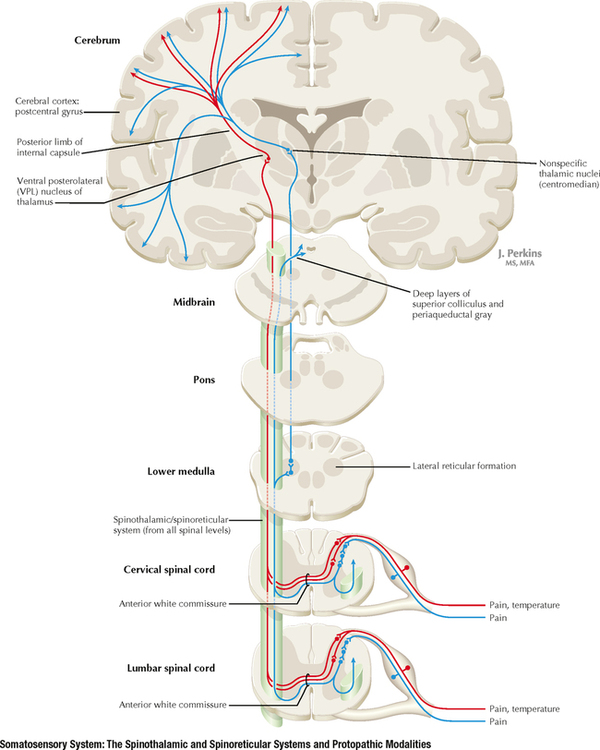
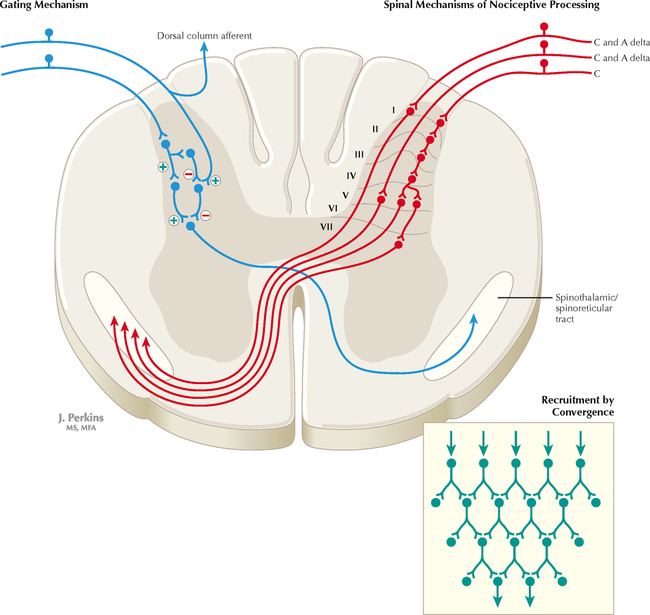
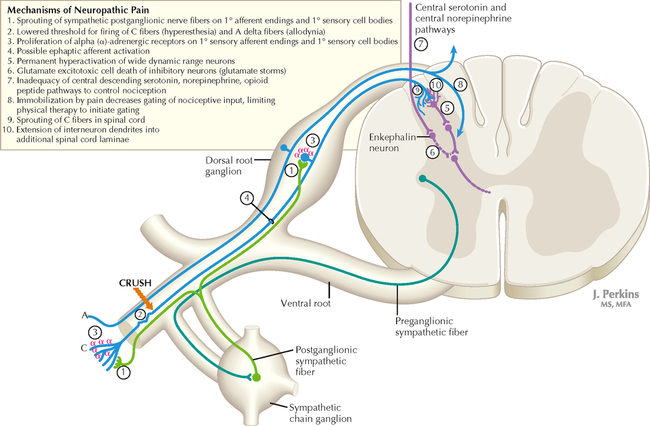
14 SENSORY SYSTEMS Somatosensory Systems 14.1. Somatosensory Afferents to the Spinal Cord 14.2. Spinal Somatic Reflex Actions and Pathways 14.3. Somatosensory System: Spinocerebellar Pathways 14.4. Somatosensory System: The Dorsal Column System and Epicritic Modalities 14.5. Somatosensory System: The Spinothalamic and Spinoreticular Systems and Protopathic Modalities 14.6. Spinothalamic and Spinoreticular Nociceptive Processing in the Spinal Cord 14.7. Mechanisms of Neuropathic Pain and Sympathetically Maintained Pain 14.8. Descending Control of Ascending Somatosensory Systems Trigeminal Sensory System 14.9. Trigeminal Sensory and Associated Sensory Systems Sensory System for Taste 14.10. Anatomy of Taste Buds and Their Receptors 14.11. Taste Pathways Auditory System 14.12. Peripheral Pathways for Sound Reception 14.13. Bony and Membranous Labyrinths 14.14. VIII Nerve Innervation of Hair Cells of the Organ of Corti 14.15. Cochlear Receptors 14.16. Afferent Auditory Pathways 14.17. Centrifugal (Efferent) Auditory Pathways Vestibular System 14.18. Vestibular Receptors 14.19. Vestibular Pathways 14.20. Nystagmus Visual System 14.21. Anatomy of the Eye 14.22. Anterior and Posterior Chambers of the Eye 14.23. The Retina: Retinal Layers 14.24. The Retina: Photoreceptors 14.25. The Retina: Optic Nerve 14.26. Arteries and Veins of the Eye 14.27. Anatomy and Relationships of the Optic Chiasm 14.28. Visual Pathways: Retinal Projections to the Thalamus, Hypothalamus, and Brain Stem 14.29. Visual Pathway: The Retino-Geniculo-Calcarine Pathway 14.30. Visual Pathways in the Parietal and Temporal Lobes 14.31. Visual System Lesions SOMATOSENSORY SYSTEMS 14.1 SOMATOSENSORY AFFERENTS TO THE SPINAL CORD Unmyelinated (UNM) and small myelinated (M) axons that convey nociception and temperature sensation terminate in lamina I and V (origin of the spinothalamic tract). Other UNM axons terminate in the dorsal horn, from which neurons for polysynaptic reflexes and for the spinoreticular system originate. M axons for touch and pressure terminate in the dorsal horn, from which additional reflex connections, spinothalamic projections, and supplementary epicritic projections to the dorsal column (DC) nuclei originate. M axons also project directly into fasciculi gracilis and cuneatus, destined for nuclei gracilis and cuneatus; these lemniscal pathways process epicritic information for conscious interpretation. M proprioceptive axons (Ia afferents) terminate directly on lower motor neurons (LMNs) and on the Ia interneuronal pool. Additional M axons terminate in the dorsal horn on neurons of origin for the spinocerebellar tracts. CLINICAL POINT Primary afferents include both epicritic afferents (mainly larger diameter M axons that convey fine, discriminative touch, vibratory sensation, and joint position sense) and protopathic afferents (mainly small M or UNM axons that convey mainly nociceptive information and temperature sensation). These axons can be affected differentially in neuropathies. Some peripheral neuropathies can affect all modalities, leading to a total loss of sensation; other peripheral neuropathies affect selected populations of axons and their related modalities. Selective loss of protopathic modalities may occur in leprosy, in amyloid neuropathy, and in some cases of diabetic neuropathy, leading to insensitivity to pain and temperature. Selective loss of epicritic sensation may occur in some distal symmetrical polyneuropathies, neuropathy with vitamin B12 deficiency, Guillain-Barré syndrome, and others, accompanied by paresthesias (numbness and tingling, “pins and needles,” abnormal sensations), dysesthesias (disagreeable or abnormal sensations in the absence of stimulation), hyperesthesia (increased sensation with stimulation), or hypesthesia (diminished sensation with stimulation). Some neuropathic conditions also are accompanied by allodynia (pain evoked by normally nonpainful stimuli) and burning, stabbing, radiating pain. Peripheral neuropathies that affect larger diameter, M axons often can also affect the motor axons, leading to weakness and hyporeflexia or areflexia. Some small fiber neuropathies, especially diabetic neuropathies, may affect small autonomic axons to bowel, bladder, reproductive organs, and peripheral blood vessels, leading to orthostatic hypotension, bladder dysfunction, chronic gastrointestinal problems, or erectile dysfunction. CLINICAL POINT The monosynaptic reflex (the muscle stretch reflex) is tested in a clinical neurological examination. Specific muscle tendons are tapped, with the expected result of contraction of the homonymous muscle (e.g., tapping of the patellar tendon resulting in contraction of the ipsilateral quadriceps muscle). The muscle stretch reflexes routinely tested in a neurological examination include the biceps reflex, triceps reflex, brachioradialis reflex, patellar (knee-jerk) reflex, and the ankle-jerk reflex on both sides. The reflexes are graded on a numerical scale ranging from hyporeflexic to normoreflexic to hyperreflexic; normal physiological reflexes may vary in responsiveness, so the result of reflex testing must be considered in conjunction with other clinical signs and symptoms. For example, hyperreflexia in a pathological state such as stroke or spinal cord injury may be accompanied by hypertonia of the affected muscle, spasticity, abnormal reflexes (extensor plantar response), and repetitive hyperreflexic responses (clonus). In contrast, hyporeflexia or areflexia accompanying peripheral neuropathy may be accompanied by muscle weakness and flaccidity and diminished sensation of epicritic modalities, protopathic modalities, or both. More formal testing of reflexic responses can be done with electromyography and conduction velocity studies. 14.2 SPINAL SOMATIC REFLEX ACTIONS AND PATHWAYS A, Presynaptic inhibition. Some interneurons synapse on the terminal arborizations of other axons, as in the case of some afferent pools associated with muscle stretch reflexes. These axoaxonic contacts permit the modulation of neurotransmitter release from the second (target) axon terminal by depolarization of the terminal membrane, altering the influx of Ca++. B, Muscle stretch reflex. In the muscle stretch reflex, Ia afferents excite the homonymous LMN pool directly and inhibit the antagonist LMN pool reciprocally via Ia inhibitory interneurons. C, Recurrent inhibition. Some interneurons receive recurrent collaterals from axons (e.g., LMN axons) and project back onto the dendrites or cell body of origin of that axon, usually inhibiting that neuron. This process can help to regulate the excitability and timing of excitation of the target neurons. Collaterals of LMN axons excite Renshaw cells (large interneurons), which inhibit the LMN of origin as well as LMNs projecting to synergistic muscles. Renshaw inhibition permits wiping the slate clean, after original excitation, of pools of LMNs, requiring additional incoming stimulation in order to excite these LMNs again. D, Golgi tendon organ reflex. Ib axons from Golgi tendon organs in muscle tendons terminate on pools of interneurons that inhibit LMNs to the homonymous muscle disynaptically and excite LMNs to the antagonist muscle reciprocally. The action of this reflex as a protective mechanism to prevent damage to a muscle during generation of maximal tension on the tendon is seen in attempted passive stretch of a spastic muscle; the resultant inhibition of the homonymous LMN pool is called a clasp-knife reflex. E, Flexor withdrawal reflex. A flexor reflex (also called a withdrawal reflex or a nociceptive reflex) occurs when afferents derived from a noxious stimulus terminate on pools of interneurons that excite appropriate pools of LMNs (often flexor LMNs) to bring about a protective withdrawal from the source of the noxious stimulus. These interneurons also inhibit the antagonist LMNs through reciprocal inhibition. Flexor reflexes can extend throughout the spinal cord, as happens when one touches a hot stove with a finger; the result is the removal of the entire arm, or even the entire body, away from the source of heat. These flexor reflexes may involve both sides of spinal cord. F, Renshaw cell bias. Some reflex responses such as Renshaw reflexes (see part C) may result in the distribution of influence (bias) in a manner that favors a particular type of action. Renshaw cells receive inputs from axon collaterals of both flexor and extensor LMNs, but their projections are directed mainly toward the inhibition of extensor LMNs (and through reciprocal inhibition with the excitation of flexor LMNs). Thus, the Renshaw cell response favors flexor movements and helps to inhibit extensor movements. 14.3 SOMATOSENSORY SYSTEM: SPINOCEREBELLAR PATHWAYS Proprioceptive primary somatosensory axons from joints, tendons, and ligaments (represented in this figure by Ib afferents from Golgi tendon organs) terminate on neurons of origin (border cells, dorsal horn) of the ventral spinocerebellar tract (VSCT) and the rostral spinocerebellar tract (RSCT) from the lower and upper body, respectively (level T6 is the cut-off point). Proprioceptive primary somatosensory axons from muscle spindles (represented in this figure by Ia afferents) terminate on neurons of origin (Clarke’s nucleus, lateral {external} cuneate nucleus of the medulla) of the dorsal spinocerebellar tract (DSCT) and the cuneocerebellar tract from the lower and upper body, respectively (level T6 is the cut-off point). The DSCT, RSCT, and cuneocerebellar tracts remain ipsilateral. The VSCT crosses twice, once in the anterior white commissure of the spinal cord and again in the cerebellum. CLINICAL POINT The dorsal and ventral spinocerebellar pathways travel in a conspicuous site at the lateral edge of the lateral funiculus throughout most of its length; these pathways are vulnerable to lesions that impinge on this zone of the spinal cord. They include tumors, radiculopathies with accompanying myelopathies, combined-system degeneration, demyelinating diseases, vascular infarcts in the anterior circulation of the cord, Brown-Séquard lesions, and other pathologies. Such a lesion, if superficial in the lateral funiculus, results in ipsilateral ataxia, dysmetria, clumsiness, and mild hypotonia, with impaired ability to perform heel-to-shin testing and tandem walking. However, lesions of the lateral funiculus often also involve the descending upper motor axons of the lateral corticospinal tract and the rubrospinal tract. Lesions that involve these tracts cause ipsilateral spastic hemiparesis or monoparesis, depending on the level of the lesion. The resulting spastic weakness, hyperreflexia, and hypertonus predominate in the clinical picture, thus masking the spinocerebellar symptomatology. Thus, an initial picture of spinocerebellar damage may give way to a progressive picture of spastic paresis on the same side. 14.4 SOMATOSENSORY SYSTEM: THE DORSAL COLUMN SYSTEM AND EPICRITIC MODALITIES Primary somatosensory myelinated axons conveying fine, discriminative touch, pressure, vibratory sensation, and consciousness of joint position project directly into the DC system (fasciculus gracilis for lower body, below T6, and fasciculus cuneatus for upper body, T6 and above), where they are topographically organized. They terminate in nuclei gracilis and cuneatus, respectively, from which the medial lemniscus originates. This tract crosses (decussates) in the medulla and projects to the ventroposterolateral (VPL) nucleus of the thalamus. Axons of neurons in the VPL nucleus terminate in the primary sensory cortex topographically. The entire DC/medial lemniscal system is topographically organized; the lower body is represented medially in the primary somatosensory cortex, and the upper body (and face from trigeminal projections) is represented laterally. This representation is sometimes drawn proportionally (the resultant figure is called a homunculus); information from the fingers and hands has far greater representation in the cerebral cortex than information from the back. The spinocervical system is a small supplement to the DC system. Primary afferent projections terminate in the medial part of the dorsal horn; these neurons project to the lateral cervical nucleus (in C1 and C2 only). This nucleus contributes additional crossed axons with polysynaptic mechanoreceptive information. 14.5 SOMATOSENSORY SYSTEM: THE SPINOTHALAMIC AND SPINORETICULAR SYSTEMS AND PROTOPATHIC MODALITIES Primary somatosensory unmyelinated (C fibers) and small myelinated (A delta fibers) that convey nociceptive information (fast, localizing pain), temperature sensation, and light, moving touch terminate on neurons in lamina I and V. These dorsal horn neurons send crossed axons into the spinothalamic tract, projecting to neurons in the VPL nucleus of the thalamus (red). This pool of neurons in the VPL nucleus is different from the pool receiving input from nuclei gracilis and cuneatus from the DC system. These thalamic neurons in the VPL nucleus project to the second somatosensory cortex (SII) as well as to the primary sensory cortex. Primary sensory C fibers also terminate in the dorsal horn and contribute to a large, cascading network for bilateral projections into the spinoreticular tract (blue). This system ends mainly in the reticular formation, from which polysynaptic projections lead to nonspecific, medial dorsal, and anterior thalamic nuclei. Some spinoreticular fibers also terminate in the deeper layers of the superior colliculus (spinotectal pathway) and in the periaqueductal gray. Cortical regions such as the cingulate, insular, and prefrontal regions then process and interpret nociceptive information related to slow, agonizing, excruciating pain. CLINICAL POINT The spinothalamic tract conveys lemniscal information from primary afferents for nociception and temperature sensation to secondary sensory neurons in lamina I and V of the dorsal horn of the spinal cord. These dorsal horn neurons then project contralateral spinothalamic tract axons to the VPL nucleus of the thalamus, which in turn sends some information about “fast pain” (not outlasting the duration of the stimulus) to sensory cortices I and II in the parietal lobe. This is the principal protopathic system tested in the neurological examination, using light pin prick and touching the body with test tubes containing water of various temperatures. This spinothalamic tract system does not convey chronic, agonizing, deep pain that characterizes many chronic diseases; such chronic “slow” pain is conveyed through a vast polysynaptic network through the dorsal horn of the spinal cord and then the lateral reticular formation of the brain. This processed information eventually reaches the nonspecific thalamic nuclei (such as the centromedian) and is conveyed to limbic structures for more subjective, interpretative aspects of pain. This latter spinoreticular network can be influenced by a host of other inputs, including the cortex, the limbic system, the descending forebrain and diencephalic systems, and collaterals of the DC system. Collaterals of the DC system can gate nociceptive processing through the dorsal horn by activating neurons that dampen transmission of information through the cascading dorsal horn network. This process is evoked in a simple fashion by light rubbing adjacent to an injured part of the body. In a more chronic fashion, DC stimulation (by a transcutaneous electrical nerve stimulation unit) can electrically activate large-diameter axons which then gate the painful stimuli bombarding the dorsal horn nociceptive axons. CLINICAL POINT The DC system consists of fasciculus gracilis (lower half of the body, with T6 cutoff) and fasciculus cuneatus (upper half of the body). These pathways consist of primary sensory axons conveying fine, discriminative touch sensation, vibratory sensation, and joint-position sense (the epicritic sensations) toward the first synapse in the secondary sensory nuclei gracilis and cuneatus in the caudal medulla. These epicritic sensations are called primary DC modalities, the basic information coded mainly by large-diameter myelinated axons. Additional DC modalities are sometimes tested if the primary modalities are intact, including two-point discrimination, stereognosis (knowing what an object is just by touch), and graphesthesia (interpreting a number drawn into the palm of the hand). These are considered cortical modalities of the DC system; they require that the primary DC modalities be intact and also require the ability of the sensory cortices to interpret the information conveyed and to draw conclusions about that information. If the primary modalities are impaired, there is no reason to attempt to test the cortical modalities that depend on unimpaired primary modalities. Pure lesions of the DC system do not entirely eliminate the primary epicritic modalities, they just remove some interpretive capabilities; such a patient may realize that a vibratory stimulus is being applied to the upper extremity but may be unable to distinguish vibratory stimuli of different frequencies. The dorsal portion of the lateral funiculus carries additional epicritic information to the DC nuclei from the spinal cord dorsal horn. A lesion of both the DC and the dorsal portion of the lateral funiculus results in total loss of epicritic sensation on the affected side. 14.6 SPINOTHALAMIC AND SPINORETICULAR NOCICEPTIVE PROCESSING IN THE SPINAL CORD Primary afferents (C and A delta fibers) conveying fast, localized pain and temperature sensation terminate in laminae I and V of the dorsal horn of the spinal cord, from which the crossed spinothalamic axons originate. Unmyelinated primary afferents (C fibers) also terminate on neurons in the dorsal horn, from which a cascading system involving recruitment, convergence, and polysynaptic interconnections originates. This system contributes to the spinoreticular tract (mainly crossed, but some are uncrossed), which projects into the RF and continues polysynaptically to nonspecific, medial dorsal and anterior thalamic nuclei. This system contributes to perception of excruciating pain and its emotional connotation via cortical regions such as the cingulate, insular, and prefrontal cortices. The gating mechanism, shown on the left, allows primary DC axon collaterals to dampen pain processing in the dorsal horn via inhibitory interneuronal connections that inhibit the flow of information through the cascading dorsal horn system that contributes to the spinoreticular pathway. 14.7 MECHANISMS OF NEUROPATHIC PAIN AND SYMPATHETICALLY MAINTAINED PAIN The cascading dorsal horn system receives primary afferent C fibers of nociceptive origin and projects into the spinoreticular system for the conscious interpretation of excruciating pain and neuropathic pain, shown in this illustration. Connections from the sympathetic nervous system can innervate terminals and cell bodies of primary nociceptive neurons directly. In neuropathic pain syndromes such as complex regional pain syndrome (CRPS), formerly called reflex sympathetic dystrophy (RSD), sympathetic postganglionic neurons may activate receptors on greatly sensitized primary afferent nerve terminals and cell bodies, either directly (on synapses) or indirectly (through secretion of norepinephrine into the blood); such activation may exacerbate the perception of the neuropathic pain. Multiple mechanisms are thought to contribute to sensitization of pain-related neurons and presence of chronic, agonizing neuropathic pain in CRPS and related syndromes. These mechanisms are noted in this illustration as numbered sites. Descending central noradrenergic and serotonergic projections are thought to play an important modulatory role in the processing of neuropathic and non-neuropathic pain. CLINICAL POINT Only gold members can continue reading. Log In or Register to continue Share this:Click to share on Twitter (Opens in new window)Click to share on Facebook (Opens in new window) Related Related posts: VENTRICLES AND THE CEREBROSPINAL FLUID SPINAL CORD MOTOR SYSTEMS AUTONOMIC-HYPOTHALAMIC-LIMBIC SYSTEMS TELENCEPHALON PERIPHERAL NERVOUS SYSTEM Stay updated, free articles. Join our Telegram channel Join Tags: Netters Atlas of Neuroscience with STUDENT CONSULT Online Access Jun 4, 2016 | Posted by admin in NEUROLOGY | Comments Off on SENSORY SYSTEMS Full access? Get Clinical Tree
14 SENSORY SYSTEMS Somatosensory Systems 14.1. Somatosensory Afferents to the Spinal Cord 14.2. Spinal Somatic Reflex Actions and Pathways 14.3. Somatosensory System: Spinocerebellar Pathways 14.4. Somatosensory System: The Dorsal Column System and Epicritic Modalities 14.5. Somatosensory System: The Spinothalamic and Spinoreticular Systems and Protopathic Modalities 14.6. Spinothalamic and Spinoreticular Nociceptive Processing in the Spinal Cord 14.7. Mechanisms of Neuropathic Pain and Sympathetically Maintained Pain 14.8. Descending Control of Ascending Somatosensory Systems Trigeminal Sensory System 14.9. Trigeminal Sensory and Associated Sensory Systems Sensory System for Taste 14.10. Anatomy of Taste Buds and Their Receptors 14.11. Taste Pathways Auditory System 14.12. Peripheral Pathways for Sound Reception 14.13. Bony and Membranous Labyrinths 14.14. VIII Nerve Innervation of Hair Cells of the Organ of Corti 14.15. Cochlear Receptors 14.16. Afferent Auditory Pathways 14.17. Centrifugal (Efferent) Auditory Pathways Vestibular System 14.18. Vestibular Receptors 14.19. Vestibular Pathways 14.20. Nystagmus Visual System 14.21. Anatomy of the Eye 14.22. Anterior and Posterior Chambers of the Eye 14.23. The Retina: Retinal Layers 14.24. The Retina: Photoreceptors 14.25. The Retina: Optic Nerve 14.26. Arteries and Veins of the Eye 14.27. Anatomy and Relationships of the Optic Chiasm 14.28. Visual Pathways: Retinal Projections to the Thalamus, Hypothalamus, and Brain Stem 14.29. Visual Pathway: The Retino-Geniculo-Calcarine Pathway 14.30. Visual Pathways in the Parietal and Temporal Lobes 14.31. Visual System Lesions SOMATOSENSORY SYSTEMS 14.1 SOMATOSENSORY AFFERENTS TO THE SPINAL CORD Unmyelinated (UNM) and small myelinated (M) axons that convey nociception and temperature sensation terminate in lamina I and V (origin of the spinothalamic tract). Other UNM axons terminate in the dorsal horn, from which neurons for polysynaptic reflexes and for the spinoreticular system originate. M axons for touch and pressure terminate in the dorsal horn, from which additional reflex connections, spinothalamic projections, and supplementary epicritic projections to the dorsal column (DC) nuclei originate. M axons also project directly into fasciculi gracilis and cuneatus, destined for nuclei gracilis and cuneatus; these lemniscal pathways process epicritic information for conscious interpretation. M proprioceptive axons (Ia afferents) terminate directly on lower motor neurons (LMNs) and on the Ia interneuronal pool. Additional M axons terminate in the dorsal horn on neurons of origin for the spinocerebellar tracts. CLINICAL POINT Primary afferents include both epicritic afferents (mainly larger diameter M axons that convey fine, discriminative touch, vibratory sensation, and joint position sense) and protopathic afferents (mainly small M or UNM axons that convey mainly nociceptive information and temperature sensation). These axons can be affected differentially in neuropathies. Some peripheral neuropathies can affect all modalities, leading to a total loss of sensation; other peripheral neuropathies affect selected populations of axons and their related modalities. Selective loss of protopathic modalities may occur in leprosy, in amyloid neuropathy, and in some cases of diabetic neuropathy, leading to insensitivity to pain and temperature. Selective loss of epicritic sensation may occur in some distal symmetrical polyneuropathies, neuropathy with vitamin B12 deficiency, Guillain-Barré syndrome, and others, accompanied by paresthesias (numbness and tingling, “pins and needles,” abnormal sensations), dysesthesias (disagreeable or abnormal sensations in the absence of stimulation), hyperesthesia (increased sensation with stimulation), or hypesthesia (diminished sensation with stimulation). Some neuropathic conditions also are accompanied by allodynia (pain evoked by normally nonpainful stimuli) and burning, stabbing, radiating pain. Peripheral neuropathies that affect larger diameter, M axons often can also affect the motor axons, leading to weakness and hyporeflexia or areflexia. Some small fiber neuropathies, especially diabetic neuropathies, may affect small autonomic axons to bowel, bladder, reproductive organs, and peripheral blood vessels, leading to orthostatic hypotension, bladder dysfunction, chronic gastrointestinal problems, or erectile dysfunction. CLINICAL POINT The monosynaptic reflex (the muscle stretch reflex) is tested in a clinical neurological examination. Specific muscle tendons are tapped, with the expected result of contraction of the homonymous muscle (e.g., tapping of the patellar tendon resulting in contraction of the ipsilateral quadriceps muscle). The muscle stretch reflexes routinely tested in a neurological examination include the biceps reflex, triceps reflex, brachioradialis reflex, patellar (knee-jerk) reflex, and the ankle-jerk reflex on both sides. The reflexes are graded on a numerical scale ranging from hyporeflexic to normoreflexic to hyperreflexic; normal physiological reflexes may vary in responsiveness, so the result of reflex testing must be considered in conjunction with other clinical signs and symptoms. For example, hyperreflexia in a pathological state such as stroke or spinal cord injury may be accompanied by hypertonia of the affected muscle, spasticity, abnormal reflexes (extensor plantar response), and repetitive hyperreflexic responses (clonus). In contrast, hyporeflexia or areflexia accompanying peripheral neuropathy may be accompanied by muscle weakness and flaccidity and diminished sensation of epicritic modalities, protopathic modalities, or both. More formal testing of reflexic responses can be done with electromyography and conduction velocity studies. 14.2 SPINAL SOMATIC REFLEX ACTIONS AND PATHWAYS A, Presynaptic inhibition. Some interneurons synapse on the terminal arborizations of other axons, as in the case of some afferent pools associated with muscle stretch reflexes. These axoaxonic contacts permit the modulation of neurotransmitter release from the second (target) axon terminal by depolarization of the terminal membrane, altering the influx of Ca++. B, Muscle stretch reflex. In the muscle stretch reflex, Ia afferents excite the homonymous LMN pool directly and inhibit the antagonist LMN pool reciprocally via Ia inhibitory interneurons. C, Recurrent inhibition. Some interneurons receive recurrent collaterals from axons (e.g., LMN axons) and project back onto the dendrites or cell body of origin of that axon, usually inhibiting that neuron. This process can help to regulate the excitability and timing of excitation of the target neurons. Collaterals of LMN axons excite Renshaw cells (large interneurons), which inhibit the LMN of origin as well as LMNs projecting to synergistic muscles. Renshaw inhibition permits wiping the slate clean, after original excitation, of pools of LMNs, requiring additional incoming stimulation in order to excite these LMNs again. D, Golgi tendon organ reflex. Ib axons from Golgi tendon organs in muscle tendons terminate on pools of interneurons that inhibit LMNs to the homonymous muscle disynaptically and excite LMNs to the antagonist muscle reciprocally. The action of this reflex as a protective mechanism to prevent damage to a muscle during generation of maximal tension on the tendon is seen in attempted passive stretch of a spastic muscle; the resultant inhibition of the homonymous LMN pool is called a clasp-knife reflex. E, Flexor withdrawal reflex. A flexor reflex (also called a withdrawal reflex or a nociceptive reflex) occurs when afferents derived from a noxious stimulus terminate on pools of interneurons that excite appropriate pools of LMNs (often flexor LMNs) to bring about a protective withdrawal from the source of the noxious stimulus. These interneurons also inhibit the antagonist LMNs through reciprocal inhibition. Flexor reflexes can extend throughout the spinal cord, as happens when one touches a hot stove with a finger; the result is the removal of the entire arm, or even the entire body, away from the source of heat. These flexor reflexes may involve both sides of spinal cord. F, Renshaw cell bias. Some reflex responses such as Renshaw reflexes (see part C) may result in the distribution of influence (bias) in a manner that favors a particular type of action. Renshaw cells receive inputs from axon collaterals of both flexor and extensor LMNs, but their projections are directed mainly toward the inhibition of extensor LMNs (and through reciprocal inhibition with the excitation of flexor LMNs). Thus, the Renshaw cell response favors flexor movements and helps to inhibit extensor movements. 14.3 SOMATOSENSORY SYSTEM: SPINOCEREBELLAR PATHWAYS Proprioceptive primary somatosensory axons from joints, tendons, and ligaments (represented in this figure by Ib afferents from Golgi tendon organs) terminate on neurons of origin (border cells, dorsal horn) of the ventral spinocerebellar tract (VSCT) and the rostral spinocerebellar tract (RSCT) from the lower and upper body, respectively (level T6 is the cut-off point). Proprioceptive primary somatosensory axons from muscle spindles (represented in this figure by Ia afferents) terminate on neurons of origin (Clarke’s nucleus, lateral {external} cuneate nucleus of the medulla) of the dorsal spinocerebellar tract (DSCT) and the cuneocerebellar tract from the lower and upper body, respectively (level T6 is the cut-off point). The DSCT, RSCT, and cuneocerebellar tracts remain ipsilateral. The VSCT crosses twice, once in the anterior white commissure of the spinal cord and again in the cerebellum. CLINICAL POINT The dorsal and ventral spinocerebellar pathways travel in a conspicuous site at the lateral edge of the lateral funiculus throughout most of its length; these pathways are vulnerable to lesions that impinge on this zone of the spinal cord. They include tumors, radiculopathies with accompanying myelopathies, combined-system degeneration, demyelinating diseases, vascular infarcts in the anterior circulation of the cord, Brown-Séquard lesions, and other pathologies. Such a lesion, if superficial in the lateral funiculus, results in ipsilateral ataxia, dysmetria, clumsiness, and mild hypotonia, with impaired ability to perform heel-to-shin testing and tandem walking. However, lesions of the lateral funiculus often also involve the descending upper motor axons of the lateral corticospinal tract and the rubrospinal tract. Lesions that involve these tracts cause ipsilateral spastic hemiparesis or monoparesis, depending on the level of the lesion. The resulting spastic weakness, hyperreflexia, and hypertonus predominate in the clinical picture, thus masking the spinocerebellar symptomatology. Thus, an initial picture of spinocerebellar damage may give way to a progressive picture of spastic paresis on the same side. 14.4 SOMATOSENSORY SYSTEM: THE DORSAL COLUMN SYSTEM AND EPICRITIC MODALITIES Primary somatosensory myelinated axons conveying fine, discriminative touch, pressure, vibratory sensation, and consciousness of joint position project directly into the DC system (fasciculus gracilis for lower body, below T6, and fasciculus cuneatus for upper body, T6 and above), where they are topographically organized. They terminate in nuclei gracilis and cuneatus, respectively, from which the medial lemniscus originates. This tract crosses (decussates) in the medulla and projects to the ventroposterolateral (VPL) nucleus of the thalamus. Axons of neurons in the VPL nucleus terminate in the primary sensory cortex topographically. The entire DC/medial lemniscal system is topographically organized; the lower body is represented medially in the primary somatosensory cortex, and the upper body (and face from trigeminal projections) is represented laterally. This representation is sometimes drawn proportionally (the resultant figure is called a homunculus); information from the fingers and hands has far greater representation in the cerebral cortex than information from the back. The spinocervical system is a small supplement to the DC system. Primary afferent projections terminate in the medial part of the dorsal horn; these neurons project to the lateral cervical nucleus (in C1 and C2 only). This nucleus contributes additional crossed axons with polysynaptic mechanoreceptive information. 14.5 SOMATOSENSORY SYSTEM: THE SPINOTHALAMIC AND SPINORETICULAR SYSTEMS AND PROTOPATHIC MODALITIES Primary somatosensory unmyelinated (C fibers) and small myelinated (A delta fibers) that convey nociceptive information (fast, localizing pain), temperature sensation, and light, moving touch terminate on neurons in lamina I and V. These dorsal horn neurons send crossed axons into the spinothalamic tract, projecting to neurons in the VPL nucleus of the thalamus (red). This pool of neurons in the VPL nucleus is different from the pool receiving input from nuclei gracilis and cuneatus from the DC system. These thalamic neurons in the VPL nucleus project to the second somatosensory cortex (SII) as well as to the primary sensory cortex. Primary sensory C fibers also terminate in the dorsal horn and contribute to a large, cascading network for bilateral projections into the spinoreticular tract (blue). This system ends mainly in the reticular formation, from which polysynaptic projections lead to nonspecific, medial dorsal, and anterior thalamic nuclei. Some spinoreticular fibers also terminate in the deeper layers of the superior colliculus (spinotectal pathway) and in the periaqueductal gray. Cortical regions such as the cingulate, insular, and prefrontal regions then process and interpret nociceptive information related to slow, agonizing, excruciating pain. CLINICAL POINT The spinothalamic tract conveys lemniscal information from primary afferents for nociception and temperature sensation to secondary sensory neurons in lamina I and V of the dorsal horn of the spinal cord. These dorsal horn neurons then project contralateral spinothalamic tract axons to the VPL nucleus of the thalamus, which in turn sends some information about “fast pain” (not outlasting the duration of the stimulus) to sensory cortices I and II in the parietal lobe. This is the principal protopathic system tested in the neurological examination, using light pin prick and touching the body with test tubes containing water of various temperatures. This spinothalamic tract system does not convey chronic, agonizing, deep pain that characterizes many chronic diseases; such chronic “slow” pain is conveyed through a vast polysynaptic network through the dorsal horn of the spinal cord and then the lateral reticular formation of the brain. This processed information eventually reaches the nonspecific thalamic nuclei (such as the centromedian) and is conveyed to limbic structures for more subjective, interpretative aspects of pain. This latter spinoreticular network can be influenced by a host of other inputs, including the cortex, the limbic system, the descending forebrain and diencephalic systems, and collaterals of the DC system. Collaterals of the DC system can gate nociceptive processing through the dorsal horn by activating neurons that dampen transmission of information through the cascading dorsal horn network. This process is evoked in a simple fashion by light rubbing adjacent to an injured part of the body. In a more chronic fashion, DC stimulation (by a transcutaneous electrical nerve stimulation unit) can electrically activate large-diameter axons which then gate the painful stimuli bombarding the dorsal horn nociceptive axons. CLINICAL POINT The DC system consists of fasciculus gracilis (lower half of the body, with T6 cutoff) and fasciculus cuneatus (upper half of the body). These pathways consist of primary sensory axons conveying fine, discriminative touch sensation, vibratory sensation, and joint-position sense (the epicritic sensations) toward the first synapse in the secondary sensory nuclei gracilis and cuneatus in the caudal medulla. These epicritic sensations are called primary DC modalities, the basic information coded mainly by large-diameter myelinated axons. Additional DC modalities are sometimes tested if the primary modalities are intact, including two-point discrimination, stereognosis (knowing what an object is just by touch), and graphesthesia (interpreting a number drawn into the palm of the hand). These are considered cortical modalities of the DC system; they require that the primary DC modalities be intact and also require the ability of the sensory cortices to interpret the information conveyed and to draw conclusions about that information. If the primary modalities are impaired, there is no reason to attempt to test the cortical modalities that depend on unimpaired primary modalities. Pure lesions of the DC system do not entirely eliminate the primary epicritic modalities, they just remove some interpretive capabilities; such a patient may realize that a vibratory stimulus is being applied to the upper extremity but may be unable to distinguish vibratory stimuli of different frequencies. The dorsal portion of the lateral funiculus carries additional epicritic information to the DC nuclei from the spinal cord dorsal horn. A lesion of both the DC and the dorsal portion of the lateral funiculus results in total loss of epicritic sensation on the affected side. 14.6 SPINOTHALAMIC AND SPINORETICULAR NOCICEPTIVE PROCESSING IN THE SPINAL CORD Primary afferents (C and A delta fibers) conveying fast, localized pain and temperature sensation terminate in laminae I and V of the dorsal horn of the spinal cord, from which the crossed spinothalamic axons originate. Unmyelinated primary afferents (C fibers) also terminate on neurons in the dorsal horn, from which a cascading system involving recruitment, convergence, and polysynaptic interconnections originates. This system contributes to the spinoreticular tract (mainly crossed, but some are uncrossed), which projects into the RF and continues polysynaptically to nonspecific, medial dorsal and anterior thalamic nuclei. This system contributes to perception of excruciating pain and its emotional connotation via cortical regions such as the cingulate, insular, and prefrontal cortices. The gating mechanism, shown on the left, allows primary DC axon collaterals to dampen pain processing in the dorsal horn via inhibitory interneuronal connections that inhibit the flow of information through the cascading dorsal horn system that contributes to the spinoreticular pathway. 14.7 MECHANISMS OF NEUROPATHIC PAIN AND SYMPATHETICALLY MAINTAINED PAIN The cascading dorsal horn system receives primary afferent C fibers of nociceptive origin and projects into the spinoreticular system for the conscious interpretation of excruciating pain and neuropathic pain, shown in this illustration. Connections from the sympathetic nervous system can innervate terminals and cell bodies of primary nociceptive neurons directly. In neuropathic pain syndromes such as complex regional pain syndrome (CRPS), formerly called reflex sympathetic dystrophy (RSD), sympathetic postganglionic neurons may activate receptors on greatly sensitized primary afferent nerve terminals and cell bodies, either directly (on synapses) or indirectly (through secretion of norepinephrine into the blood); such activation may exacerbate the perception of the neuropathic pain. Multiple mechanisms are thought to contribute to sensitization of pain-related neurons and presence of chronic, agonizing neuropathic pain in CRPS and related syndromes. These mechanisms are noted in this illustration as numbered sites. Descending central noradrenergic and serotonergic projections are thought to play an important modulatory role in the processing of neuropathic and non-neuropathic pain. CLINICAL POINT Only gold members can continue reading. Log In or Register to continue Share this:Click to share on Twitter (Opens in new window)Click to share on Facebook (Opens in new window) Related Related posts: VENTRICLES AND THE CEREBROSPINAL FLUID SPINAL CORD MOTOR SYSTEMS AUTONOMIC-HYPOTHALAMIC-LIMBIC SYSTEMS TELENCEPHALON PERIPHERAL NERVOUS SYSTEM Stay updated, free articles. Join our Telegram channel Join Tags: Netters Atlas of Neuroscience with STUDENT CONSULT Online Access Jun 4, 2016 | Posted by admin in NEUROLOGY | Comments Off on SENSORY SYSTEMS Full access? Get Clinical Tree